









专利摘要
本发明涉及岷江百合类萌发素蛋白基因LrGLP1的应用,LrGLP1基因的核苷酸序列如SEQIDNO:1所述,编码类萌发素蛋白,本发明通过功能基因组学相关技术研究证实LrGLP1基因具有提高植物抗真菌侵染的功能。将本发明抗真菌LrGLP1基因构建到植物表达载体上并转入烟草中过量表达,转基因烟草植株具有很强的体外抗真菌活性。LrGLP1超表达的转基因烟草对链格孢、核盘菌以及串珠状赤霉菌的生长具有明显的抑制作用。
权利要求
1.一种岷江百合类萌发素蛋白基因LrGLP1在提高烟草对链格孢、核盘菌、串珠状赤霉菌抗性中的应用。
2.根据权利要求1所述的岷江百合类萌发素蛋白基因LrGLP1的应用,其特征在于提高烟草的真菌抗性的具体操作如下:
(1)将岷江百合类萌发素蛋白基因LrGLP1与植物超表达载体pCAMBIA2300S连接,构建植物超表达载体;
(2)将上述构建的重组载体通过根癌农杆菌介导转入烟草中;
(3)以重组载体T-DNA上具有的抗性标记来筛选转化子,并通过聚合酶链式反应筛选获得真正的转基因植株,接种特定病原真菌,分析转基因烟草蛋白对真菌生长的抑制活性,最后筛选出对真菌抗性明显增强的转基因植株。
说明书
技术领域
本发明涉及分子生物学以及基因工程相关技术研究领域,特别是具有抗真菌活性的岷江百合类萌发素蛋白基因LrGLP1的应用。
背景技术
植物病害是指植物在生物因素的影响下发生的一系列生化、生理甚至形态上的病变,阻碍正常的生长、发育,进而严重危害农业生产。病原真菌是引起植物病害的主要病原物。真菌病害发病率极高,且大规模发生时会致使农作物减产甚至死亡。传统对于真菌病害的预防和处理主要是筛选抗性强的品种、及时清理病害植株及果实、使用农药等,这些措施有的周期长,有的效果差,有的还会给环境和人畜的生命健康带来危害,因此不能彻底防治真菌病害。随着重组DNA技术的创立和快速发展,利用基因工程技术来培育新的对真菌病害有很强抗性的植物品种已经成为解决植物真菌病害的新思路,有望从根本上解决真菌病害问题。
与萌发素(germin)一样,类萌发素蛋白(Germin-like proteins, GLPs)最初是在小麦中发现(Dunwell J M, Gibbings J G, Mahmood T, et al. Germin and germin-like proteins: evolution, structure, and function. Critical Reviews in Plant Sciences, 2008, 27(5): 342-375)。在结构上,两者都是由两个外显子编码形成的稳定低聚物,都含有β-折叠桶状结构域,且都含有一个“cupin”蛋白结构域,共同属于一个功能多样化的家族,即cupin超家族。在GLPs的二级结构中包含一个特征结构域称为“germin box”,结构域中包含两个特定基序,分别为:germin motif 1-G(x)5HxH(x)3,4E(x)6G和germin motif 2-G(x)5PxG(x)2H(x)3N (Breen J, Bellgard M. Germin-like proteins (GLPs) in cereal genomes: gene clustering and dynamic roles in plant defence. Functional & Integrative Genomics, 2010, 10(4): 463-476)。GLPs为多功能蛋白,主要以酶、受体和结构蛋白的形式参与多种生理生化过程。其中,酶主要包括超氧化物歧化酶(superoxide dismutase,SOD)、草酸盐氧化酶(oxalate oxidase,OXO)、ADP葡萄糖焦磷酸酶/磷酸二酯酶(ADP glucose pyrophosphatase/phosphodiesterase,AGPPase),受体如雄激素结合蛋白(androgen binding protein,ABP19/20)激素受体、Rhicadhesins受体等。
目前根据功能的差异将植物的GLPs分为3个亚类:第1亚类为“true germin”,主要包括小麦、大麦的萌发素及一些其他谷类植物的GLPs,它们具有SOD和OXO的双重活性;第2亚类的GLPs主要是来自于除小麦和大麦之外的其他禾谷类、裸子植物和茄科植物等,这一亚类主要与植物耐氧化胁迫有直接关系,并与锰超氧化物歧化酶(MnSOD)极其相似;第3亚类的GLPs主要包括与生长素代谢相关的一些调节蛋白,它们与植物的生理节律和花期诱导功能有关(Khuri S, Bakker F T, Dunwell J M. Phylogeny, function, and evolution of the cupins, a structurally conserved, functionally diverse superfamily of proteins. Molecular Biology and Evolution, 2001, 18(4): 593-605)。Woo等人(2000)研究一个大麦germin蛋白的空间结构时发现,每个gemin蛋白以单体的形式相互结合形成二聚体,再进一步以“二聚体的三聚体”的方式形成极为稳定的六聚体(Woo EJ, Dunwell JM, Goodenough PW, et al. Germin is a manganese containing homohexamer with oxalate oxidase and superoxide dismutase activities. Nature Structural Biology, 2000, 7: 1036-1040)。与MnSOD类似,Gemin单体中存在相应的配体并可以结合锰离子,这就从一方面解释了GLP蛋白具有SOD活性的原因。在GLPs中包含两个半胱氨酸残基,可以形成二硫键,维持蛋白质空间结构的稳定性。
Godfrey等人在葡萄(Vitis vinifera)中已经发现7个GLPs基因,这些GLPs的ORF所编码蛋白长度从207-205个氨基酸残基不等,并且包含不同的GLP特征结构域(Godfrey D, Able A J, Dry I B. Induction of a grapevine germin-like protein (VvGLP3) gene is closely linked to the site of Erysiphe necator infection: a possible role in defense. Molecular Plant-Microbe Interactions, 2007, 20(9): 1112-1125)。VvGLPs的表达在不同组织和不同发育阶段差异较大。VvGLP2和VvGLP6在葡萄叶和浆果(包括果皮和果肉)中有表达;VvGLP1和VvGLP5分别只在根和成熟之前的浆果中表达;VvGLP3、VvGLP4、VvGLP7则在所有的组织中都表达。白粉病菌(Erysiphe necator)侵染迅速诱导浆果和叶片中VvGLP3和VvGLP4的表达,相对而言,白粉病菌侵染的浆果中VvGLP5和发病叶中VvGLP6的表达量都显著减少,VvGLP2的表达却基本不受白粉病菌侵染的影响。VvGLP3的表达受E. necator侵染的诱导,并具有SOD活性。从甜菜(Beta vulgaris)克隆的GLP基因BvGLP-1,在线虫(Heterodera schachtii)入侵后转录水平上调,将BvGLP-1转入拟南芥中过表达,转基因株系抑制真菌菌丝在体内外的生长扩散,进而增强植株对病原真菌Verticillium longisporum和Rhizoctonia solani的抗性,但是不会影响有益真菌在其体内的生长(Knecht K, Seyffarth M, Desel C, et al. Expression of BvGLP-1 encoding a germin-like protein from sugar beet in Arabidopsis thaliana leads to resistance against phytopathogenic fungi. Molecular Plant-Microbe Interactions, 2010, 23(4): 446-457)。
GLPs在植物的抗病防卫反应中有着极为重要的作用。通过RNA干扰使水稻(Oryza sativa)类萌发素蛋白基因OsGLP1沉默,OsGLP1功能缺失植株的高度只有大约正常植株的一半,谷粒也不饱满,同时转基因植株对真菌病害的敏感度增加,如纹枯病和稻瘟病(Banerjee J, Maiti M K. Functional role of rice germin-like protein1 in regulation of plant height and disease resistance. Biochemical and Biophysical Research Communications, 2010, 394(1): 178-183)。BnGLP3同时具有OXO和SOD活性,还可以在氧爆发时水解活性氧生成H2O2,降低植株对病原菌的敏感性,且在油菜(Brassica napus)受核盘菌(Sclerotinia sclerotiorum)侵染6h后表达量迅速增加(Rietz S, Bernsdorff F E M, Cai D. Members of the germin-like protein family in Brassica napus are candidates for the initiation of an oxidative burst that impedes pathogenesis of Sclerotinia sclerotiorum. Journal of experimental botany, 2012, 63(15): 5507-5519)。
本发明中类萌发素蛋白基因LrGLP1来自岷江百合(Lilium regale Wilson)。岷江百合又名王百合,多年生草本植物,我国的百合特有种。仅分布于四川西岷江流域海拔800~2700m的河谷到山腰的岩石缝中,对尖孢镰刀菌等真菌具有很强的抗性。
发明内容
本发明的目的是提供一种从岷江百合中克隆获得具有抗真菌活性的类萌发素蛋白的全长基因LrGLP1,LrGLP1的核苷酸序列如SEQ ID NO:1所示,该基因全长为908bp,包含一个654bp的开放阅读框、59bp的5’非翻译区(untranslated regions, UTR)及195bp的3’UTR,编码如SEQ ID NO:2所示氨基酸序列的蛋白质。
本发明所述类萌发素蛋白基因LrGLP1的编码区是序列表SEQ ID NO:1中第60-713位所示的核苷酸序列。
本发明分离克隆岷江百合的一个抗真菌相关基因的完整cDNA片段,通过根癌农杆菌(Agrobacterium tumefaciens)介导将目的基因转入受体植物中过量表达,并通过进一步实验验证该基因是否具有抗真菌的活性,为后期利用该基因改良烟草及其他植物抵御真菌病害的能力奠定基础。发明人将这个基因命名为LrGLP1。
GLP是PR家族中的一类胞外糖蛋白,在植物的生长发育以及对生物和非生物胁迫的防御反应中有重要的作用。GLPs具有OXO、SOD、AGPPase中的至少一种活性。在真菌病害入侵时,SOD将氧爆发时形成的活性氧转化为过氧化氢和氧分子,降低过多活性氧对植株形成的伤害;OXO可以催化草酸产生二氧化碳和过氧化氢;两种反应所产生的过氧化氢可以通过纤维素交联作用增强细胞壁的结构,并催化细胞壁的氧化交联形成乳突,延缓和阻止病原菌的侵入和扩散,以保护细胞免受再次感染。此外过氧化氢还是植物体内重要的抗病防卫反应相关的信号分子,可以激活并诱导体内大量抗病基因的表达,激活体内的抗病防卫反应。
本发明涉及分离包含LrGLP1的DNA片段并鉴定其功能,具有该基因片段的植物在一定程度上具有抵抗特定真菌入侵的表型。其中所述DNA片段如序列表SEQ ID所示,对该基因进行分析,表明LrGLP1全长cDNA为908bp,包含一个654bp的开放阅读框、59bp的5’UTR及195bp的3’UTR,其中ORF编码一个具有217氨基酸的蛋白质。LrGLP1的核苷酸序列与岷江百合LrGLP2有97%的相似性。编码蛋白具有类萌发素蛋白的特征结构域,germin motif 1和germin motif 2。BLASTp检索结果表明LrGLP1编码的蛋白质与岷江百合LrGLP2的相似性最高,与山羊草(Aegilops tauschii)小麦(Triticum uratu) 中几个GLPs的相似性约为64%~65%,这表明其属于岷江百合中的类萌发素蛋白。超表达序列表SEQ ID:1所示序列可以增强烟草对链格孢(Alternaria alternata)、核盘菌(Sclerotinia sclerotiorum)以及串珠状赤霉菌(Gibberella moniliformis)的抗性。
上述LrGLP1基因可以应用于提高烟草的抗真菌特性,具体操作如下:
(1)采用扩增LrGLP1的特异引物,从接种尖孢镰刀菌后的岷江百合根中提取总RNA,通过逆转录-聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR)扩增出LrGLP1的全长编码区,然后将其连接到pMD-18T载体上,经测序获得具有目的基因的克隆。
(2)用限制性内切酶BamHI和EcoRI酶切pMD18-T-LrGLP1载体和植物表达载体pCAMBIA2300S,通过胶回收得到目的基因片段和载体大片段。再将所获得LrGLP1基因片段与pCAMBIA2300S载体片段连接,构建植物超表达载体。之后将所构建的重组载体通过根癌农杆菌介导转入烟草中表达。
(3)以重组载体T-DNA上具有的抗性标记筛选转化子,并通过PCR以及RT-PCR检测得到真正的转基因植株,分析转基因植株对于病原真菌的抗性,最后筛选出对真菌抗性明显增强的转基因植株。
本发明为提高植物对真菌病害的抗性提供了一种新的方法,通过基因工程手段培育抗病植物可以克服传统育种的不足,不仅育种周期缩短,而且操作简单,容易获得高抗材料。本发明中来自岷江百合的LrGLP1基因能增强植物对几种病原真菌的抗性,将该基因导入烟草中,可以产生具有真菌抗性的新品种和新材料。利用基因工程技术培育抗性植物品种和材料具有明显的优势和不可取代的重要性。它不仅可以为大规模生产作物、花卉等提供方便,减少化学农药的使用,还可以为农业生产节约成本、减少环境污染,因此本发明具有广阔的市场应用前景。
附图说明
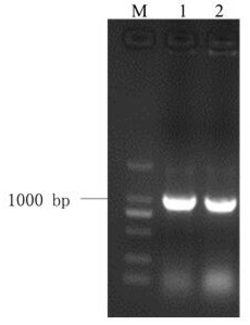
图1是本发明中部分LrGLP1转基因烟草基因组DNA的PCR检测结果,其中Marker:DL2000 DNA Marker (大连宝生物),由2,000bp、1,000bp、750bp、500bp、250bp以及100bp六条DNA片段组成;正对照:质粒pMD18-T-LrGLP1为模板的PCR反应;WT:非转基因烟草(野生型)总DNA为模板进行的PCR。
图2是本发明中部分阳性LrGLP1转基因烟草中LrGLP1转录水平的表达分析结果图,其中Marker:DL2000 DNA Marker(大连宝生物);WT:非转基因烟草总RNA逆转录cDNA为模板的PCR产物;正对照:质粒pMD18-T- LrGLP1为模板的PCR产物。
图3是本发明中LrGLP1转基因烟草体外抗真菌活性的抑菌效果图;其中a、b、c图示中的真菌分别是链格孢、核盘菌、串珠状赤霉菌;WT为野生型烟草的总蛋白;CK为空白对照,即无蛋白对照(用于提取蛋白的缓冲液)。
具体实施方式
下面通过附图和实施例对本发明进一步说明,但本发明保护范围不局限于所述内容,本实施例中方法如无特殊说明的均按常规方法操作,所用试剂如无特殊说明的采用常规试剂或按常规方法配置的试剂。
实施例1:LrGLP1全长cDNA克隆以及序列分析
用尖孢镰刀菌接种岷江百合,用接种后12 h的根提取总RNA,用液氮将处理过的岷江百合的根研磨成粉末,然后转入离心管中,采用异硫氰酸胍法提取总RNA。采用逆转录酶M-MLV (promega)以总RNA为模板合成cDNA第一链,反应体系和操作过程为:取5 μg total RNA,依次加入50 ng oligo (dT),2 μL dNTP Mix (2.5mM each),用DEPC水将反应体积补齐至14.5 μL;混匀后,70℃加热变性5 min后迅速在冰上冷却5 min,然后依次加入4 μL 5×First-stand buffer、0.5 μL RNasin (200U)、1 μL M-MLV (200U),混匀并简短离心,42℃温浴1.5 h,取出后70℃加热10 min,终止反应。cDNA第一链合成后置于-20℃保存备用。
以合成的第一链cDNA为模板,扩增目的基因LrGLP1,所用上下游引物序列分别为5’ACGCACATATGGCTACCCACTACT3’及5’ CGACTAGAACTGAGCCTGGAGCC3’。采用AdvantageTM 2 PCR Enzyme (Clontech)扩增出目的基因。PCR反应条件:95℃ 1 min;94℃ 30 s,61℃ 30 s,72℃ 1 min,28个循环;72℃ 5 min。反应体系(20 μL)为1 μL cDNA、2 μL 10×Advantage 2 PCR Buffer、1.8 μL dNTP Mix (10mM each)、0.2 μL 正向引物(10 μM)、0.2 μL 反向引物(10 μM)、0.2 μL Advantage 2 PCR Polymerase Mix、14.6 μL PCR-Grade water。PCR结束后,取8 μL进行琼脂糖凝胶电泳,用以检测扩增产物的特异性以及大小。
所得到PCR产物只有一条DNA带,故直接对PCR产物进行TA克隆,使用的试剂盒为pMD18-T vector kit (大连宝生物),反应体系和操作过程为:取1.5 μL PCR产物,依次加入1 μL pMD18-T vector (50 ng/μL)和2.5 μL 2×Ligation solution I,混匀后置于16℃过夜反应。通过热激转化法将连接产物转入大肠杆菌DH5α感受态中。用含有氨苄青霉素(ampicillin,Amp)的LB固体培养基筛选阳性克隆。挑选若干个单菌落,摇菌后用扩增LrGLP1的特异引物检测多克隆位点插入LrGLP1的克隆。将得到的阳性克隆进行测序,最终获得的LrGLP1全长cDNA为908 bp,通过NCBI ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析发现其包含一个654bp的开放读码框(见序列表)。LrGLP1编码一个含217个氨基酸的蛋白质LrGLP1,其分子量约为23.36 KDa,等电点为5.60。借助生物信息学软件SignalP 4.1分析LrGLP1编码的蛋白序列,检测其是否具有N端信号肽。结果显示在LrGLP1中存在信号肽,这就表明LrGLP1为一种分泌蛋白。
实施例2:植物超表达载体构建
采用SanPrep柱式质粒DNA小量抽提试剂盒(上海生工)提取插入LrGLP1的大肠杆菌质粒pMD18-T-LrGLP1以及植物表达载体pCAMBIA2300S质粒,取1 μL用于琼脂糖凝胶电泳以检测所提取质粒的完整性及浓度高低。用限制性内切酶EcoRI (TaKaRa)和BamHI (TaKaRa)分别对质粒pMD18-T-LrGLP1和pCAMBIA2300S进行双酶切(100 μL体系),反应体系和操作过程为:分别取20 μL pMD18-T-LrGLP1和pCAMBIA2300S质粒、依次加入10 μL 10×K buffer、5 μL EcoRI、5 μL BamHI、60 μL ddH2O,混匀后短时离心,置于37℃过夜反应。将所有酶切产物进行琼脂糖凝胶电泳,然后使用SanPrep柱式DNA胶回收试剂盒 (上海生工)对LrGLP1片段和pCAMBIA2300s载体大片段分别进行胶回收,取1 μL回收产物通过琼脂糖凝胶电泳检测回收片段的大小以及浓度,置于-20℃保存备用。
利用T4 DNA Ligase (TaKaRa),将回收的LrGLP1DNA片段和pCAMBIA2300S载体片段连接起来,反应体系(20 μL)和操作过程为:取10 μL LrGLP1DNA片段依次加入2 μL pCAMBIA2300S载体DNA、2 μL 10×T4 DNA Ligase Buffer、1 μL T4 DNA Ligase、5 μL ddH2O,混匀后短时离心,然后16℃水浴过夜反应。接着采用热激转化法将连接产物转入大肠杆菌DH5α中,用含有50mg/L卡那霉素(kanamycin,Km)的固体培养基筛选阳性克隆。挑选单菌落摇菌,以菌液为模板用扩增LrGLP1的特异引物进行PCR,挑选出LrGLP1与pCAMBIA2300S成功连接的克隆,并向检测得到的阳性菌株中加入甘油并置于-80℃保存备用。
采用SanPrep柱式质粒抽提试剂盒(上海生工)提取并纯化上述大肠杆菌DH5α中的pCAMBIA2300S-LrGLP1质粒。随后用液氮冻融法将上述构建的植物表达载体pCAMBIA2300S-LrGLP1转入所制备的根癌农杆菌LBA4404感受态细胞中。操作步骤为:取2 μg pCAMBIA2300S-LrGLP1质粒加入含有200 μL感受态细胞的离心管中,轻轻混匀后冰浴5 min,随后转入液氮中冷冻1 min,然后迅速置于37℃水浴5 min,再冰浴2 min,之后加入500 μL LB液体培养基于28℃振荡培养4 h。将活化后的农杆菌涂于含有50 mg/L Km的LB固体培养基上,28℃倒置培养。挑选单菌落摇菌,再用扩增LrGLP1的特异性引物进行PCR反应,检测pCAMBIA2300S-LrGLP1是否转入农杆菌中。对于阳性克隆,加入甘油后置于-80℃保存备用。
实施例3:农杆菌介导的植物遗传转化以及转基因植物筛选
本实验的转基因受体是烟草(Nicotiana tabacum L.)。将烟草种子用75%的酒精浸泡30 s,用无菌水洗涤后用0.1%的HgCl2浸泡8 min,然后再用无菌水洗涤若干次,播种于1/2 MS培养基上,28℃暗培养5-8 d,发芽后转至光照培养箱(25℃,16h/d光照),以后每月用MS培养基继代一次。
从-80℃冰箱中取出保存的含有pCAMBIA2300S-LrGLP1质粒的农杆菌LBA4404菌种,取20 μL接种于5 mL含有50 mg/L Km和20 mg/L利福平的LB液体培养基中,28℃培养至培养基浑浊。吸取1 mL浑浊的菌液至含有50 mg/L Km的LB固体培养基上,28℃培养48 h。随后将LB固体培养基上的农杆菌刮下适量接种于附加有20 mg/L的乙酰丁香酮的MGL液体培养基中,28℃振荡培养5-8 h以活化农杆菌。
取无菌烟草苗叶子切成约1 cm2的叶盘,完全浸泡于上述含有活化农杆菌的MGL液体培养基中,25℃浸染15 min。用无菌滤纸吸干叶盘表面的菌液,将叶盘置于共培养基上,22℃无光条件下共培养2天。烟草转化的共培养基为MS+0.02 mg/L 6-BA+2.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂。
将共培养后的叶盘转到加有抗生素的MS筛选培养基中分化成苗,同时筛选转基因植株。烟草筛选培养基为MS+0.5 mg/L 6-BA+0.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂+50 mg/L Km+200 mg/L 头孢霉素(cefotaxime sodium salt,Cef);筛选培养时将培养瓶转移至光照培养箱培养(25℃,16h/d光照,8h/d黑暗)。待烟草长出芽后用含有50 mg/L Km和200 mg/L Cef的MS培养基继代培养。因烟草愈伤分化率较高,故需要对再生植株进行进一步筛选。将烟草再生苗移至含有50 mg/L Km的MS培养基上使其生根,最后选用生根较好的再生苗做进一步的检测。
采用CTAB法提取转基因烟草植株叶片的基因组DNA,取1 μL所得基因组DNA进行琼脂糖凝胶电泳检测其完整性和浓度。以转基因植株的基因组DNA为模板用LrGLP1的特异引物进行PCR反应。PCR结束后,取8 μL产物用于琼脂糖凝胶电泳以检测阳性转基因植株。部分烟草转基因植株的扩增结果如图1所示。LrGLP1转基因烟草共筛选到29株阳性转基因植株。
实施例4:转基因烟草中LrGLP1的表达分析以及转基因植株抗真菌活性分析
分别取阳性转基因植株以及非转基因烟草(野生型)的嫩叶提取总RNA,逆转录生成cDNA第一链,并以此为模板用扩增LrGLP1的特异引物进行PCR,根据PCR结果分析各转基因植株中LrGLP1转录水平的表达。总RNA提取以及RT-PCR的方法与实施例1中相同。PCR结束之后,取8 μL用于琼脂糖凝胶电泳,部分单株的检测结果如图2所示。共检测到18个转基因单株中LrGLP1在转录水平大量表达,这些单株的编号为1~18。
将实验室保存的几种真菌接种于PDA固体培养基(200 g/L马铃薯,15 g/L琼脂,20 g/L葡萄糖)上,28℃暗培养,待菌落生长至直径约为2~3cm时添加蛋白,分析转基因植株体外抗真菌活性。供试真菌共有6种:葡萄座腔菌(Botrosphaeria dothidea)、链格孢 (Alternaria alternata)、灰葡萄孢(Botrytis cinerea)、胶孢炭疽菌(Colletorichum gloeosporioides)、串珠状赤霉菌(Gibberella moniliformis)和核盘菌(Sclerotinia scleroterum)。为了防止其它杂菌污染所提取的蛋白,整个植物蛋白提取过程均是无菌操作。首先取1 g转基因烟草单株(编号分别为2、5、6、10)及野生型叶片放入研钵中,加入1 mL蛋白提取液(1M NaCl,0.1M 乙酸钠,1% PVP,pH6),充分研磨。转入1.5 mL离心管中,混匀后4℃静置过夜。4℃离心30 min (12,000 g),取上清于新的1.5 mL离心管中,并取适量用紫外分光光度仪测定总蛋白浓度。将转基因和野生型植株的总蛋白浓度调整至0.2 μg/μL,然后分别取20 μL滴于各真菌培养基的无菌滤纸上。在每个真菌的平板上除了添加不同转基因烟草植株的总蛋白,同时平行添加野生型烟草的总蛋白和空白对照(蛋白提取液)。28℃培养几天后观察各处理真菌生长的情况,并据此来评价LrGLP1转基因烟草的体外抗真菌活性。结果如图3所示,LrGLP1转基因烟草蛋白对核盘菌、链格孢以及串珠状赤霉菌的生长具有明显的抑制作用。
序列表(SEQ ID)
<110> 昆明理工大学
<120> 岷江百合类萌发素蛋白基因LrGLP1的应用
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 908
<212> DNA
<213> Lilium regaleWilson
<220>
<221> mRNA
<222> (1)..(908)
<220>
<221> 5'UTR
<222> (1)..(59)
<220>
<221> CDS
<222> (60)..(713)
<220>
<221> 3'UTR
<222> (714)..(908)
<400> 1
acatgggaag tccttcacct ttgagaaaat cgtctcgatt gctaaaatat aacgcacata 60
tggctaccca ctacttcctc tggctcgccg tccttgccct agccccctcc cattcaatgg 120
cttctgatcc tgactcactg caggatttct gtgtggctga cccaaactcc caagtgattg 180
tgaatggatt agtctgcaag gacccaaagc aagcccaacc tgctgatttt ttcttccccg 240
gactgaacca gccagggagc acaaccaacc agctggggtc taacgtcact cttgtcagcg 300
ccgcaaacct tccggggctc aacgcccttg gaatctctgt ggctcgcctg gactttgcac 360
cgtatggcct cattccccct cactaccatc cacgtgcgac cgagatcctg acattgctgg 420
agggtactct ctacgttggc ttcgtcactt ccttccccga cttccgtctc atcagcaaga 480
tcctcaaggc cggtgatgtc tttgtcttcc ccaaaggact cgtccacttc cagtttaatc 540
aggacggcac caaaccagca gtttccctgt caggcttgag cagccagaac ccaggactgg 600
tcactgtagc taatgccgtt ttcggatcta acccgcctat ctgggatgat gttctggcca 660
aggccttcca gttggacaag aagacagttg attggctcca ggctcagttc tagtcgggtt 720
gccctaatga tagcctgctg atagtagcaa aacatgtcga tgtttgtatt gactataaag 780
taataagcaa gcagatgctt tcatgattgt atgttcaatt ttccattaca taattaagtg 840
ttataatata tttaaatatg tttttttttc taattaagaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaa 908
<210> 2
<211> 217
<212> PRT
<213> Lilium regale Wilson
<400> 2
Met Ala Thr His Tyr Phe Leu Trp Leu Ala Val Leu Ala Leu Ala Pro
1 5 10 15
Ser His Ser Met Ala Ser Asp Pro Asp Ser Leu Gln Asp Phe Cys Val
20 25 30
Ala Asp Pro Asn Ser Gln Val Ile Val Asn Gly Leu Val Cys Lys Asp
35 40 45
Pro Lys Gln Ala Gln Pro Ala Asp Phe Phe Phe Pro Gly Leu Asn Gln
50 55 60
Pro Gly Ser Thr Thr Asn Gln Leu Gly Ser Asn Val Thr Leu Val Ser
65 70 75 80
Ala Ala Asn Leu Pro Gly Leu Asn Ala Leu Gly Ile Ser Val Ala Arg
85 90 95
Leu Asp Phe Ala Pro Tyr Gly Leu Ile Pro Pro His Tyr His Pro Arg
100 105 110
Ala Thr Glu Ile Leu Thr Leu Leu Glu Gly Thr Leu Tyr Val Gly Phe
115 120 125
Val Thr Ser Phe Pro Asp Phe Arg Leu Ile Ser Lys Ile Leu Lys Ala
130 135 140
Gly Asp Val Phe Val Phe Pro Lys Gly Leu Val His Phe Gln Phe Asn
145 150 155 160
Gln Asp Gly Thr Lys Pro Ala Val Ser Leu Ser Gly Leu Ser Ser Gln
165 170 175
Asn Pro Gly Leu Val Thr Val Ala Asn Ala Val Phe Gly Ser Asn Pro
180 185 190
Pro Ile Trp Asp Asp Val Leu Ala Lys Ala Phe Gln Leu Asp Lys Lys
195 200 205
Thr Val Asp Trp Leu Gln Ala Gln Phe
210 215
<210> 3
<211> 24
<212> DNA
<213> 人工序列
<400> 3
acgcacatat ggctacccac tact 24
<210> 4
<211> 23
<212> DNA
<213> 人工序列
<400> 4
cgactagaac tgagcctgga gcc 23
岷江百合类萌发素蛋白基因LrGLP1的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0