









专利摘要
本发明公开了漾濞大泡核桃病程相关蛋白10基因JsPR10-1的应用,本发明通过功能基因组学相关技术研究证实JsPR10-1基因具有提高植物抗真菌病害的功能,将本发明抗真菌基因JsPR10-1构建到植物表达载体上并转入烟草中过量表达,转基因烟草植株具有很强的体外抗真菌活性,JsPR10-1超表达的转基因烟草对胶孢炭疽菌、核盘菌、葡萄座腔菌以及串珠状赤霉菌的生长具有明显的抑制作用。
权利要求
1.漾濞大泡核桃病程相关蛋白10基因JsPR10-1在提高烟草对胶孢炭疽菌、核盘菌、葡萄座腔菌、串珠状赤霉菌抗性中的应用。
2.根据权利要求1所述的漾濞大泡核桃病程相关蛋白10基因JsPR10-1的应用,其特征在于提高烟草的真菌抗性的具体操作如下:
(1)将漾濞大泡核桃病程相关蛋白10基因JsPR10-1与植物超表达载体pCAMBIA2300S连接,构建植物超表达载体;
(2)将上述构建的重组载体通过根癌农杆菌介导转入烟草中;
(3)用卡那霉素筛选再生的转基因烟草幼苗,并通过聚合酶链式反应筛选获得阳性转基因植株,接种特定的植物病原真菌,分析转基因烟草蛋白对真菌生长的抑制活性,最后筛选出对真菌抗性明显增强的转基因植株。
说明书
技术领域
本发明涉及分子生物学以及基因工程相关研究领域,特别是具有抗真菌活性的漾濞大泡核桃病程相关蛋白10基因JsPR10-1的应用。
背景技术
21世纪,人类面临耕地减少、人口增加和人民生活水平不断提高三个不可逆转的局面。提高农作物单产是21世纪农业的重要措施,但植物病害严重影响农作物的产量和质量,其中,真菌病害是最重要的一类植物病害。传统控制植物病害的方法主要是选育抗病品种和使用化学农药,虽然取得了一些效果,但常规育种周期较长,投入的人力、物力较大,防治效果不稳定,使用农药对人畜、环境等的安全问题等缺陷,因而两者均不能从根本上解决植物病害的难题。随着分子生物学和生物技术的快速发展,利用分子生物学技术获得具有抗真菌活性的基因,并通过转基因技术培育抗病品种,是提高植物抗病能力的新方法。
植物病程相关蛋白(pathogenesis-related protein,PR)是植物在受到细菌、真菌和病毒等病原体入侵或受到非生物胁迫时产生并积累的一类蛋白质的总称,是植物参与防御反应的重要组成部分。PR蛋白最早是van Loon等于1970年在烟草花叶病毒侵染的烟草叶片中发现的 (Van Loon, L. C., Pierpoint, W. S., Boller, T. H., & Conejero, V. 1994. Recommendations for naming plant pathogenesis-related proteins. Plant Molecular Biology Reporter, 12: 245–264.)。PR蛋白在不同植物中表现为几丁质酶、葡聚糖苷酶、蛋白酶抑制剂、过氧化物酶和类核糖核酸酶等。根据其结构、亲源关系和生物活性,PR蛋白主要分为17个亚类:PR1~PR17 (Van Loon, L. C., Rep, M., & Pieterse, C. M. J. 2006. Significance of inducible defense-related proteins in infected plants. Annu Rev Phytopathol, 44: 135–162.),广泛存在于单子叶和双子叶植物中。其中一类具有核酸酶活性的蛋白定义为第10类PR蛋白(简称PR10),引起很多研究者的兴趣。
与大多数胞外PR蛋白不同,PR10是一类胞内蛋白。PR10蛋白具有与核酸酶相似的结构,分子量为16~19 kDa,等电点偏酸性,大部分特异定位在细胞质中,具有体外核酸酶活性和抗菌活性。对PR10基因结构分析表明,PR10基因编码的氨基酸中都具有一个高度保守的GXGGXG(X是任意氨基酸)序列模型,被称为“P-loop”结构域。“P-loop”是一类广泛存在于磷酸化激酶和核酸结合蛋白中的结构域,并且其磷酸化可能与PR10的核糖核酸酶活性有关(Bantignies, B., Séguin, J., Muzac, I., Dédaldéchamp, F., Gulick, P., & Ibrahim, R. 2000. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots. Plant molecular biology, 42: 871–881.)。
研究表明,多种外源激发因子(细菌、真菌、病毒等病原物)和内源激发因子(水杨酸、乙烯、过氧化氢等)均能诱导PR10的基因表达。与此同时,PR10也受多种非生物胁迫的诱导,如盐、热、冷、紫外光等。Chadha和Das研究发现花生AhPR-10基因受病原菌的诱导,其融合蛋白在体外能显著地抑制Fusarium oxysporum和Rhizoctonia solani的生长(Chadha, P., & Das, R. H. 2006. A pathogenesis related protein, AhPR10 from peanut: an insight of its mode of antifungal activity. Planta, 225: 213–222.)。在水稻中,茉莉酸诱导RSOsPR10的表达上调(Hashimoto, M., Kisseleva, L., Sawa, S., Furukawa, T., Komatsu, S., & Koshiba, T. 2004. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway. Plant and cell physiology, 45: 550–559.)。Park等研究辣椒CaPR-10基因时发现,其在高盐处理后30 min开始积累,6 h后表达最强(Park, C. J., Kim, K. J., Shin, R., Park, J. M., Shin, Y. C., & Paek, K. H. 2004. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. The Plant Journal, 37: 186–198.)。虽然很多PR10蛋白都是在病原物侵染或者某些信号分子(如水杨酸、乙烯等)的诱导下生成和大量积累的,但也有一些研究表明,PR10在正常生长的植株中存在组成型表达,如玉米的ZmPR-10和ZmPR-10.1,它们都在根中高水平表达,但ZmPR-10在各个组织的表达量明显高于ZmPR-10.1 (Xie, Y. R., Chen, Z. Y., Brown, R. L., & Bhatnagar, D. 2010. Expression and functional characterization of two pathogenesis-related protein 10 genes from Zea mays. Journal of plant physiology, 167: 121–130.)。
PR10广泛应答生物胁迫,具有抗菌活性。来自高丽参(Panax ginseng C. A. Meyer)的PgPR10-1基因在拟南芥中异源表达,接种丁香假单胞菌(Pseudomonas syringe)、尖孢镰刀菌(F. oxysporum)和灰霉病菌(Botrytis cinerea)4天后,统计转基因和野生型植株感病叶片数,结果表明,与野生型植株相比,转基因拟南芥增强了对以上三种病原菌的抗性(Lee, O. R., Kim, Y. J., Balusamy, S. R. D., Khorolragchaa, A., Sathiyaraj, G., Kim, M. K., & Yang, D. C. 2012. Expression of the ginseng PgPR10-1 in Arabidopsis confers resistance against fungal and bacterial infection. Gene, 506: 85–92.)。从感染黄曲霉(Aspergillus flavus)的花生(Arachis hypogaea L.)中克隆得到ARAhPR10,并转入花生中高效表达,ARAhPR10转基因花生种子抗A. hypogaea能力明显提高(Xie, C., Wen, S., Liu, H., Chen, X., Li, H., Hong, Y., & Liang, X. 2013. Overexpression of ARAhPR10, a member of the PR10 family, decreases levels of Aspergillus flavus infection in peanut seeds. American Journal of Plant Sciences, 4: 602.)。在Escherichia coli BL21中表达的棉花(Gossypium barbadense) GbPR10-1,体外抑菌实验显示,GbPR10-1蛋白可以抑制Verticillium dahliae菌丝的生长(Chen, L., Sun, N., Wang, J., Ling, H., Zhang, L., & Zuo, K. 2013. Functional analysis of a wilt fungus inducible PR10-1 gene from cotton. American Journal of Plant Sciences, 4: 417.)。
本发明中病程相关蛋白基因JsPR10-1来自漾濞大泡核桃(Juglans sigillata Dode)。漾濞大泡核桃是云南主要的本地核桃之一,具有果大、壳薄、仁厚、味香、含油率高等优点,对由胶孢炭疽菌引起的病害具有较强的抗性。
发明内容
本发明的目的是提供漾濞大泡核桃病程相关蛋白10基因JsPR10-1的新用途,即在提高烟草对胶孢炭疽菌、核盘菌、葡萄座腔菌、串珠状赤霉菌抗性中的应用。
本发明从漾濞大泡核桃中克隆获得的具有抗真菌活性的病程相关蛋白10的全长基因JsPR10-1,JsPR10-1的核苷酸序列如SEQ ID NO:1所示,该基因全长为776 bp,包含一个483 bp的开放阅读框、74 bp的5′非翻译区(untranslated region, UTR)及219 bp的3′UTR,编码如SEQ ID NO:2所示氨基酸序列的蛋白质。
本发明所述病程相关蛋白10基因JsPR10-1的编码区是序列表SEQ ID NO:1中第75-557位所示的核苷酸序列。
本发明分离克隆漾濞大泡核桃的一个抗真菌相关基因的完整cDNA片段,通过根癌农杆菌(Agrobacterium tumefaciens)介导将目的基因转入受体植物中过量表达,并通过进一步实验验证该基因是否具有抗真菌的活性,为后期利用该基因改良烟草及其他植物抵御真菌病害的能力奠定基础。发明人将这个基因命名为JsPR10-1。
特异定位于细胞质的PR10蛋白具有与核酸酶相似的结构,是分子量为16~19kDa的酸性蛋白,具有核酸酶活性和体外抑菌活性,能响应生物和非生物胁迫,是植物防御系统中重要的组成部分。
本发明涉及分离包含JsPR10-1的DNA片段并鉴定其功能,具有该基因片段的植物在一定程度上具有抵抗特定真菌入侵的表型。其中所述DNA片段如序列表所示,对该基因进行分析,表明JsPR10-1全长cDNA为776 bp,包含一个483 bp的开放阅读框、74 bp的5′UTR及219bp的3′UTR,其中ORF编码一个具有160个氨基酸的蛋白质。BLASTn分析结果显示漾濞大泡核桃JsPR10-1的第82~259位核苷酸序列与桦树PR10 (Betula nigra, EU526188.1)的第1~178位核苷酸序列具有83%的相似性。JsPR10-1中存在一个高度保守的氨基酸序列GXGGXG (X为任意氨基酸),被称为“P-loop”基序。蛋白质同源性分析表明JsPR10-1编码的蛋白质序列与栎树Betv 1 allergen (Quercus suber, AFF59689.1)、欧洲山毛榉stress and pathogenesis-related protein (Fagus sylvatica, CAA10235.1)以及欧洲板栗ypr10 (Castanea sativa, CAD10374.1)高度同源,相似性分别为83%、79%和77%,这表明其属于漾濞大泡核桃中的病程相关蛋白10。超表达序列表SEQ ID NO:1所示序列可以增强烟草对胶孢炭疽菌(Colletotrichum gloeosporioides)、核盘菌(Sclerotinia sclerotiorum)、葡萄座腔菌(Botrosphaeria dothidea)以及串珠状赤霉菌(Gibberella moniliformis)的抗性。
上述JsPR10-1基因可以应用于提高烟草的抗真菌特性,具体操作如下:
(1)采用扩增JsPR10-1的特异引物,从接种胶孢炭疽菌后的漾濞大泡核桃叶中提取总RNA,通过逆转录-聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR)扩增出JsPR10-1的全长编码区,然后将其连接到pMD-18T载体上,经测序获得具有目的基因的克隆。
(2)用限制性内切酶PstⅠ和EcoRI酶切pMD18-T-JsPR10-1载体和植物表达载体pCAMBIA2300S,通过胶回收得到目的基因片段和载体大片段。再将所获得JsPR10-1基因片段与pCAMBIA2300S载体片段连接,构建植物超表达载体。之后将所构建的重组载体通过根癌农杆菌介导转入烟草中表达。
(3)以重组载体T-DNA上具有的抗性标记筛选转化子,并通过PCR以及RT-PCR检测得到真正的转基因植株,分析转基因植株对于植物病原真菌的抗性,最后筛选出对真菌抗性明显增强的转基因植株。
本发明为提高植物对真菌病害的抗性提供了一种新的方法,通过基因工程手段培育抗病植物可以克服传统育种的不足,不仅育种周期缩短,而且操作简单,容易获得高抗材料。本发明中来自漾濞大泡核桃的JsPR10-1基因能增强植物对几种病原真菌的抗性,将该基因导入烟草中,可以产生具有真菌抗性的新品种和新材料。利用基因工程技术培育抗性植物品种和材料具有明显的优势和不可取代的重要性。它不仅可以为大规模生产作物、花卉等提供方便,减少化学农药的使用,还可以为农业生产节约成本、减少环境污染,因此本发明具有广阔的市场应用前景。
附图说明
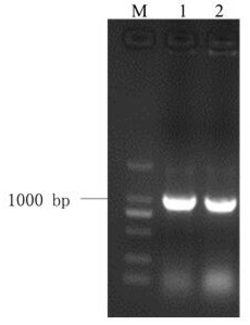
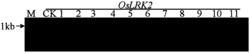
图1是本发明中部分JsPR10-1转基因烟草基因组DNA的PCR检测结果,其中Marker:DL2000 DNA Marker (大连宝生物),由2,000 bp、1,000 bp、750 bp、500 bp、250 bp以及100 bp六条DNA片段组成;正对照:质粒pMD18-T-JsPR10-1为模板的PCR反应;WT:非转基因烟草(野生型)总DNA为模板进行的PCR;
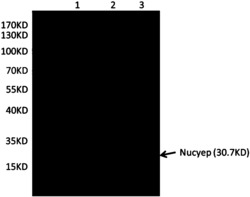
图2是本发明中部分阳性JsPR10-1转基因烟草中JsPR10-1转录水平的表达分析结果图,其中Marker:DL2000 DNA Marker (大连宝生物);WT:非转基因烟草总RNA逆转录cDNA为模板的PCR产物;正对照:质粒pMD18-T-JsPR10-1为模板的PCR产物;
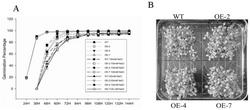
图3是本发明中JsPR10-1转基因烟草体外抗真菌活性的抑菌效果图;其中a、b、c、d、图示中的真菌分别是核盘菌、胶孢炭疽菌、串珠状赤霉菌、葡萄座腔菌;WT为野生型烟草的总蛋白;CK为空白对照,即无蛋白对照(用于提取蛋白的缓冲液)。
具体实施方式
下面通过附图和实施例对本发明进一步说明,但本发明保护范围不局限于所述内容,本实施例中方法如无特殊说明的均按常规方法操作,所用试剂如无特殊说明的采用常规试剂或按常规方法配置的试剂。
实施例1:JsPR10-1全长cDNA克隆以及序列分析
用胶孢炭疽菌接种漾濞大泡核桃的嫩叶,用接种后4 h的叶提取总RNA,用液氮将处理过的漾濞大泡核桃的叶研磨成粉末,然后转入离心管中,采用异硫氰酸胍法提取总RNA。采用M-MLV逆转录酶(promega)以总RNA为模板合成cDNA第一链,反应体系和操作过程为:取5 μg total RNA,依次加入50 ng oligo (dT),2 μL dNTP Mix (2.5 mM each),用DEPC水将反应体积补齐至14.5 μL;混匀后,70℃加热变性5 min后迅速在冰上冷却5 min,然后依次加入4 μL 5×First-stand buffer、0.5 μL RNasin (200 U)、1 μL M-MLV (200 U),混匀并简短离心,42℃温浴1.5 h,取出后70℃加热10 min,终止反应。cDNA第一链合成后置于-20℃保存备用。
以合成的第一链cDNA为模板,扩增目的基因JsPR10-1,所用上下游引物序列分别为 -ACTCCTTTTCCTTTTACCACTTCCA- 及 -AACAACTCGCAGGGTCGCAGA- 。采用AdvantageTM 2 PCR Enzyme (Clontech)扩增出目的基因。PCR反应条件:95℃ 1 min;94℃ 30 s,62℃ 30 s,72℃ 45 s,32个循环;72℃ 5 min。反应体系(20μL)为1 μL cDNA、2 μL 10×Advantage 2 PCR Buffer、1.8 μL dNTP Mix (10mM each)、0.2 μL 正向引物(10 μM)、0.2 μL 反向引物(10 μM)、0.2 μL Advantage 2 PCR Polymerase Mix、14.6 μL PCR-Grade water。PCR结束后,取8 μL进行琼脂糖凝胶电泳,用以检测扩增产物的特异性以及大小。
所得到PCR产物只有一条DNA带,故直接对PCR产物进行TA克隆,使用的试剂盒为pMD18-T vector kit (大连宝生物),反应体系和操作过程为:取1.5 μL PCR产物,依次加入1 μL pMD18-T vector (50 ng/μL)和2.5 μL 2×Ligation solution I,混匀后置于16℃过夜反应。通过热激转化法将连接产物转入大肠杆菌DH5α感受态中。用含有氨苄青霉素(ampicillin,Amp)的LB固体培养基筛选阳性克隆。挑选若干个单菌落,摇菌后用扩增JsPR10-1的特异引物检测多克隆位点插入JsPR10-1的克隆。将得到的阳性克隆进行测序,最终获得的JsPR10-1全长cDNA为776 bp,通过NCBI ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析发现其包含一个483 bp的开放读码框(见序列表)。JsPR10-1编码一个含160个氨基酸的蛋白质JsPR10-1,其分子量约为17.53 KDa,等电点为5.51。借助生物信息学软件SignalP 4.1分析JsPR10-1编码的蛋白序列,检测其是否具有N端信号肽。结果显示在JsPR10-1中没有信号肽,因此推测该蛋白不是分泌蛋白。
实施例2:植物超表达载体构建
采用SanPrep柱式质粒DNA小量抽提试剂盒(上海生工)提取插入JsPR10-1的大肠杆菌质粒pMD18-T-JsPR10-1以及植物表达载体pCAMBIA2300S质粒,取1 μL用于琼脂糖凝胶电泳以检测所提取质粒的完整性及浓度高低。用限制性内切酶EcoRI (TaKaRa)和PstⅠ (TaKaRa)分别对质粒pMD18-T-JsPR10-1和pCAMBIA2300S进行双酶切(100 μL体系),反应体系和操作过程为:分别取20 μL pMD18-T-JsPR10-1和pCAMBIA2300S质粒、依次加入10 μL 10×H buffer、5 μL EcoRI、5 μL PstⅠ、60 μL ddH2O,混匀后短时离心,置于37℃过夜反应。将所有酶切产物进行琼脂糖凝胶电泳,然后使用SanPrep柱式DNA胶回收试剂盒 (上海生工)对JsPR10-1片段和pCAMBIA2300s载体大片段分别进行胶回收,取1 μL回收产物通过琼脂糖凝胶电泳检测回收片段的大小以及浓度,置于-20℃保存备用。
利用T4 DNA Ligase (TaKaRa),将回收的JsPR10-1DNA片段和pCAMBIA2300S载体片段连接起来,反应体系(20 μL)和操作过程为:取10 μL JsPR10-1DNA片段依次加入2 μL pCAMBIA2300S载体DNA、2 μL 10×T4 DNA Ligase Buffer、1 μL T4 DNA Ligase、5 μL ddH2O,混匀后短时离心,然后16℃水浴过夜反应。接着采用热激转化法将连接产物转入大肠杆菌DH5α中,用含有50 mg/L卡那霉素(kanamycin,Km)的固体培养基筛选阳性克隆。挑选单菌落摇菌,以菌液为模板用扩增JsPR10-1的特异引物进行PCR,挑选出JsPR10-1与pCAMBIA2300S成功连接的克隆,在得到的阳性菌株中加入甘油并置于-80℃保存备用。
采用SanPrep柱式质粒抽提试剂盒(上海生工)提取并纯化上述大肠杆菌DH5α中的pCAMBIA2300S-JsPR10-1质粒。随后用液氮冻融法将上述构建的植物表达载体pCAMBIA2300S-JsPR10-1转入所制备的根癌农杆菌LBA4404感受态细胞中。操作步骤为:取0.2 μg pCAMBIA2300S-JsPR10-1质粒加入含有200 μL感受态细胞的离心管中,轻轻混匀后冰浴5 min,随后转入液氮中冷冻1 min,然后迅速置于37℃水浴5 min,再冰浴2 min,之后加入500μL LB液体培养基于28℃振荡培养4 h。将活化后的农杆菌涂于含有50 mg/L Km的LB固体培养基上,28℃倒置培养。挑选单菌落摇菌,再用扩增JsPR10-1的特异性引物进行PCR反应,检测pCAMBIA2300S-JsPR10-1是否转入农杆菌中。对于阳性克隆,加入甘油后置于-80℃保存备用。
实施例3:农杆菌介导的植物遗传转化以及转基因植物筛选
本实验的转基因受体是烟草(Nicotiana tabacum L.)。将烟草种子用75%的酒精浸泡30 s,无菌水洗涤后用0.1%的HgCl2浸泡8 min,然后再用无菌水洗涤若干次,播种于1/2 MS培养基上,28℃暗培养5-8 d,发芽后转至光照培养箱(25℃,16h/d光照),以后每月用MS培养基继代一次。
从-80℃冰箱中取出保存的含有pCAMBIA2300S-JsPR10-1质粒的农杆菌LBA4404菌种,取20 μL接种于5 mL含有50 mg/L Km和20 mg/L利福平的LB液体培养基中,28℃培养至培养基浑浊。吸取1 mL浑浊的菌液至含有50 mg/L Km的LB固体培养基上,28℃培养48 h。随后将LB固体培养基上的农杆菌刮下适量接种于附加有20 mg/L的乙酰丁香酮的MGL液体培养基中,28℃振荡培养5-8 h以活化农杆菌。
取烟草无菌苗幼嫩叶切成约1 cm2的叶盘,完全浸泡于上述含有活化农杆菌的MGL液体培养基中,25℃浸染15 min。用无菌滤纸吸干叶盘表面的菌液,将叶盘置于共培养基上,22℃无光条件下共培养2天。烟草转化的共培养基为MS+0.02 mg/L 6-BA+2.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂。
将共培养后的叶盘转到加有抗生素的MS筛选培养基中分化成苗,同时筛选转基因植株。烟草筛选培养基为MS+0.5 mg/L 6-BA+0.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂+50 mg/L Km+200 mg/L 头孢霉素(cefotaxime sodium salt,Cef);筛选培养时将培养瓶转移至光照培养箱培养(25℃,16 h/d光照,8 h/d黑暗)。待烟草长出芽后用含有50 mg/L Km和200 mg/L Cef的MS培养基继代培养。因烟草愈伤分化率较高,故需要对再生植株进行进一步筛选。将烟草再生苗移至含有50 mg/L Km的MS培养基上使其生根,最后选用生根较好的再生苗做进一步的检测。
采用CTAB法提取转基因烟草植株叶片的基因组DNA,取1 μL所得基因组DNA进行琼脂糖凝胶电泳检测其完整性和浓度。以转基因植株的基因组DNA为模板用JsPR10-1的特异引物进行PCR反应。PCR结束后,取8 μL产物用于琼脂糖凝胶电泳以检测阳性转基因植株。部分烟草转基因植株的扩增结果如图1所示,JsPR10-1转基因烟草共筛选到32株阳性转基因植株。
实施例4:转基因烟草中JsPR10-1的表达分析以及转基因植株抗真菌活性分析
分别取阳性转基因植株以及非转基因烟草(野生型)的嫩叶提取总RNA,逆转录生成cDNA第一链,并以此为模板用扩增JsPR10-1的特异引物进行PCR,根据PCR结果分析各转基因植株中JsPR10-1转录水平的表达量。总RNA提取以及RT-PCR的方法与实施例1中相同。PCR结束之后,取8 μL用于琼脂糖凝胶电泳,部分单株的检测结果如图2所示,共检测到20个转基因单株中JsPR10-1在转录水平大量表达,这些单株的编号为1~20。
将实验室保存的几种真菌接种于PDA固体培养基(200 g/L马铃薯,15 g/L琼脂,20 g/L葡萄糖)上,28℃暗培养,待菌落生长至直径约为2~3 cm时添加蛋白,分析转基因植株体外抗真菌活性。供试真菌共有4种:胶孢炭疽菌(Colletorichum gloeosporioides) 、核盘菌(Sclerotinia scleroterum)、葡萄座腔菌(Botrosphaeria dothidea)和串珠状赤霉菌(Gibberella moniliformis)。为了防止其它杂菌污染所提取的蛋白,整个植物蛋白提取过程均是无菌操作。首先取1 g转基因烟草单株(编号分别为1、3、6、8)及野生型叶片放入研钵中,加入1 mL蛋白提取液(1 M NaCl,0.1 M 乙酸钠,1% PVP,pH6.0),充分研磨。转入1.5 mL离心管中,混匀后4℃静置过夜。4℃离心30 min (12,000 g),取上清于新的1.5 mL离心管中,并取适量用紫外分光光度仪测定总蛋白浓度。将转基因和野生型植株的总蛋白浓度调整至0.2 μg/μL,然后分别取20 μL滴于各真菌培养基的无菌滤纸上。在每个真菌的平板上除了添加不同转基因烟草植株的总蛋白,同时平行添加野生型烟草的总蛋白和空白对照(蛋白提取液)。28℃培养几天后观察各处理真菌生长的情况,并据此来评价JsPR10-1转基因烟草的体外抗真菌活性,结果如图3所示,JsPR10-1转基因烟草蛋白对胶孢炭疽菌、核盘菌、葡萄座腔菌以及串珠状赤霉菌的生长具有明显的抑制作用。
序列表(SEQ ID)
<110> 昆明理工大学
<120> 漾濞大泡核桃病程相关蛋白10基因JsPR10-1的应用
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 776
<212> DNA
<213> Juglans SigillataDode
<220>
<221> mRNA
<222> (1)..(776)
<220>
<221> 5'UTR
<222> (1)..(74)
<220>
<221> CDS
<222> (75)..(557)
<220>
<221> 3'UTR
<222> (558)..(776)
<400> 1
atggggacca acgcaaaaac tccccaaact ccttttcctt ttaccacttc caaagtctca 60
gttttattca tatcatgggt gttttcactt acgagactga gagcacctca gtgatccctc 120
cagctaggtt gttcaaggcc tttgttcttg atgccgataa cctcatccca aaggttgtac 180
ctcaggcagt caagagctct gaaatcattg aaggaaatgg aggtcctgga accatcaaga 240
agatcaactt tggcgaaggg agccagtaca agtatgtgaa gcacaggacc gatgcgatcg 300
acgaagcaaa cttcacatac gcctacagcg tgattgaggg agatgcactg gcagacaaga 360
tcgagaaaat ctcgtacgag acgaagatag tggcatccca tgagggagga tccattttga 420
agagcatcag ccactaccac agcaaaggcg accacgagat caaggaagaa gacgtgaagg 480
atggtaaaga gaaagcctca ggacttttca aggcagttga gggttacctg ttggcacacc 540
ctgatgctta caactaatta aattccataa atctgcgacc ctgcgagttg tttaatttgt 600
ggctttcatt tctttcgagc caagactgca tgcttttggg tttgataata aagaagaagt 660
ttgctcatgt actggtattg attatgatct gttattttgc tgtaaaactc ggctcacata 720
caaaatatat aatgaaaatc atcttgttct aaaaaaaaaa aaaaaaaaaa aaaaaa 776
<210> 2
<211> 160
<212> PRT
<213> Juglans SigillataDode
<400> 2
Met Gly Val Phe Thr Tyr Glu Thr Glu Ser Thr Ser Val Ile Pro Pro
1 5 10 15
Ala Arg Leu Phe Lys Ala Phe Val Leu Asp Ala Asp Asn Leu Ile Pro
20 25 30
Lys Val Val Pro Gln Ala Val Lys Ser Ser Glu Ile Ile Glu Gly Asn
35 40 45
Gly Gly Pro Gly Thr Ile Lys Lys Ile Asn Phe Gly Glu Gly Ser Gln
50 55 60
Tyr Lys Tyr Val Lys His Arg Thr Asp Ala Ile Asp Glu Ala Asn Phe
65 70 75 80
Thr Tyr Ala Tyr Ser Val Ile Glu Gly Asp Ala Leu Ala Asp Lys Ile
85 90 95
Glu Lys Ile Ser Tyr Glu Thr Lys Ile Val Ala Ser His Glu Gly Gly
100 105 110
Ser Ile Leu Lys Ser Ile Ser His Tyr His Ser Lys Gly Asp His Glu
115 120 125
Ile Lys Glu Glu Asp Val Lys Asp Gly Lys Glu Lys Ala Ser Gly Leu
130 135 140
Phe Lys Ala Val Glu Gly Tyr Leu Leu Ala His Pro Asp Ala Tyr Asn
145 150 155 160
<210> 3
<211> 25
<212> DNA
<213> 人工序列
<400> 3
actccttttc cttttaccac ttcca 25
<210> 4
<211> 21
<212> DNA
<213> 人工序列
<400> 4
aacaactcgc agggtcgcag a 21
漾濞大泡核桃病程相关蛋白10基因JsPR10-1的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。













动态评分
0.0