









专利摘要
本发明公开了漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1及应用,JsPRP1基因的核苷酸序列如SEQIDNO:1所示,编码富含脯氨酸蛋白,本发明通过功能基因组学相关技术研究证实JsPRP1基因具有提高植物抗真菌侵染的功能,将本发明抗真菌JsPRP1基因构建到植物表达载体上并转入烟草中过量表达,转基因烟草植株具有很强的体外抗真菌活性;JsPRP1超表达的转基因烟草对胶孢炭疽菌、核盘菌、葡萄座腔菌以及串珠状赤霉菌的生长具有明显的抑制作用。
权利要求
1.一种漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1,其特征在于:其核苷酸序列如SEQ ID NO :1 所示。
2.权利要求1 所述的漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1在提高烟草对胶孢炭疽菌、核盘菌、葡萄座腔菌、串珠状赤霉菌抗性中的应用。
3.根据权利要求2所述的漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1的应用,其特征在于提高烟草的真菌抗性的具体操作如下:
(1)将漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1与植物超表达载体pCAMBIA2300S连接,构建植物超表达载体;
(2)将上述构建的重组载体通过根癌农杆菌介导转入烟草中;
(3)以重组载体T-DNA上具有的卡那霉素抗性基因来筛选转化子,并通过聚合酶链式反应筛选获得阳性转基因植株,接种特定病原真菌,分析转基因烟草蛋白对真菌生长的抑制活性,最后筛选出对真菌抗性明显增强的转基因植株。
说明书
技术领域
本发明涉及分子生物学以及基因工程相关技术研究领域,特别是具有抗真菌活性的漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1及应用。
背景技术
植物在整个生长期内都有可能遭受病原菌的侵染。在种数繁多的植物病害中,真菌病害约占植物病害的70~80%。兴起于20世纪初期的经典遗传学使人们能够通过杂交育种成功地培育出抗病新品种,从而大幅度地提高粮食产量。然而,常规育种具有周期长、费时费力、有利变异少等缺点,不能从根本上解决植物病害难题。近年来,随着分子生物学理论和技术的不断发展,使人们不但能够从分子水平上深入认识植物与病原物相互作用的机制,而且还可以通过基因工程快速和高效地培育抗病作物新品种。
植物细胞明显区别于动物细胞的特征之一就是具有细胞壁。植物细胞壁是一个高度复杂和动态的结构,它们决定细胞的大小和形状,在增强植物细胞的机械强度、调控细胞的生长速度、物质运输、细胞间信息传递等方面起着非常重要的作用。此外,植物细胞壁还构成了阻止病原体入侵的第一道结构屏障。植物细胞壁主要由纤维素、半纤维素、果胶和蛋白质组成。蛋白质占细胞壁干重的10%左右。细胞壁蛋白质主要由结构蛋白和酶蛋白组成,结构蛋白被认为形成一个独立的决定壁结构的网络进而辅助细胞壁装配,这些蛋白质在结构上高度重复,富含特定的一两种氨基酸(许文亮, 黄耿青, 王秀兰, 汪虹, 李学宝. 一类新的编码PRPs基因的分离及其在棉花纤维等组织细胞中的表达. 生物化学与生物物理进展. 2007, 34: 509–517.)。
富含脯氨酸蛋白(proline-rich protein, PRP)是一类富含脯氨酸和羟脯氨酸的细胞壁结构蛋白,它们最先发现于胡萝卜贮藏根中,被认为是外界伤害的诱导产物(Chen J, Varner JE. Isolation and characterization of cDNA clones for carrot extensin and a proline-rich 33-kDa protein. Proceedings of the National Academy of Sciences. 1985,82: 4399–4403.)。植物富含脯氨酸蛋白可以分为三类:杂合的富含脯氨酸蛋白(HyPRPs),包含POVEKPOVXK重复序列的PRPs以及NHyPRP蛋白(Huang G, Gong S, Xu W, Li P, Zhang D, Qin L, Li W, Li X. GhHyPRP4, a cotton gene encoding putative hybrid proline-rich protein, is preferentially expressed in leaves and involved in plant response to cold stress. Acta biochimica et biophysica Sinica. 2011, 43: 519–527.)。
PRPs的表达具有组织器官特异性,并受发育阶段及环境因素的调节。Wyatt等在大豆中发现了3个编码PRPs的基因:SbPRP1、SbPRP2和SbPRP3,尽管在核苷酸和氨基酸序列上显示很高的同源性,但它们的表达特性具有较大差异。SbPRP1在大豆下胚轴的韧皮部和木质部表达,在下胚轴成熟区和伸长区的表皮细胞中也有表达;SbPRP2在下胚轴维管组织的皮层细胞中和成熟种子的内珠被中表达;SbPRP3在下胚轴快速伸长区中柱的内胚层细胞中和叶子的表皮细胞中表达 (Wyatt RE, Nagao RT, Key JL. Patterns of soybean proline-rich protein gene expression. The Plant Cell Online. 1992, 4: 99–110.)。 GmPRP2p-1062是大豆(Glycine max) GmPRP2 的启动子,在大豆的毛状根和拟南芥的根中优先表达(Chen L, Jiang B, Wu C, Sun S, Hou W, Han T. GmPRP2 promoter drives root-preferential expression in transgenic Arabidopsis and soybean hairy roots. BMC plant biology. 2014,14: 245.)。
PRPs在抵抗真菌胁迫中有重要的作用。研究表明,沉积和交联在珍珠粟(Pennisetum glaucum)植物细胞壁的PRPs对植物病原菌Sclerospora graminicola有抵抗和屏障作用(Deepak S, Shailasree S, Sujeeth N, Kini RK, Shetty SH, Mith?fer A. Purification and characterization of proline/hydroxyproline-rich glycoprotein from pearl millet coleoptiles infected with downy mildew pathogen Sclerospora graminicola. Phytochemistry. 2007, 68: 298–305.)。拟南芥中编码HyPRP的EARLI1基因具有抗真菌的功能。Northern印迹杂交结果显示,灰霉病菌(Botrytis cinerea)接种处理可以诱导拟南芥EARLI1基因的表达。采用台酚蓝染色观察真菌侵染状况,发现灰霉病菌对超表达EARLI1的植株叶片的侵染程度比野生型植株的叶片要轻,而EARLI1基因突变体植株叶片被侵染的程度较为严重(徐丹. 拟南芥EARLI1基因在生物和非生物胁迫中的抗性功能. 西北大学硕士学位论文. 2010.)。平板抑菌实验表明,重组EARLI1蛋白明显抑制了尖孢镰刀菌(Fusarium oxysporum)、灰葡萄孢(B. cinerea)分生孢子的萌发和菌丝生长(Li L, Zhang C, Xu D, Schl?ppi M, Xu ZQ. Expression of recombinant EARLI1, a hybrid proline-rich protein of Arabidopsis, in Escherichia coli and its inhibition effect to the growth of fungal pathogens and Saccharomyces cerevisiae. Gene. 2012, 506: 50–61.)。
本发明中富含脯氨酸蛋白JsPRP1来自漾濞大泡核桃(Juglans sigillata Dode)。漾濞大泡核桃是目前云南主要的核桃栽培品种,具有果大、壳薄、仁白、味香、出油率高、营养丰富等优点,并且对病原真菌胶孢炭疽菌具有较强的抗性。
发明内容
本发明的目的是提供一种从漾濞大泡核桃中克隆获得具有抗真菌活性的富含脯氨酸蛋白的全长基因JsPRP1,JsPRP1的核苷酸序列如SEQ ID NO:1所示,该基因全长为815 bp,包含一个408 bp的开放阅读框、123 bp的5′非翻译区(untranslated regions,UTR)及284 bp的3′UTR,编码如SEQ ID NO:2所示氨基酸序列的蛋白质。
本发明所述富含脯氨酸蛋白基因JsPRP1的编码区是序列表SEQ ID NO:1中第124-531位所示的核苷酸序列。
本发明分离克隆漾濞大泡核桃的一个抗真菌相关基因的完整cDNA片段,通过根癌农杆菌(Agrobacterium tumefaciens)介导将目的基因转入受体植物中过量表达,并通过进一步实验验证该基因是否具有抗真菌的活性,为后期利用该基因改良烟草及其他植物抵御真菌病害的能力奠定基础,发明人将这个基因命名为JsPRP1。
植物PRPs特异定位于细胞壁,属于细胞壁结构蛋白,但与细胞膜也有相互作用。PRPs在构建植物的细胞壁结构方面起作用,在生物和非生物胁迫下诱导表达,是植物防御系统中重要的组成成分。PRPs的表达能提高植物组织的木质化水平,增强细胞壁的机械强度,而细胞壁是病原体入侵植物的第一道物理屏障,PRPs在植物防卫反应中起到积极的作用。另外,PRPs基因的启动子片段中含有多个与脱水、病原物、高盐、低温等逆境相关的诱导元件,超表达PRP基因能提高转基因植物的抗病性。
本发明涉及分离包含JsPRP1的DNA片段并鉴定其功能,具有该基因片段的植物在一定程度上具有抵抗特定真菌入侵的表型。其中所述DNA片段如序列表所示,对该基因进行分析,表明JsPRP1全长cDNA为815 bp,包含一个408 bp的开放阅读框、123 bp的5′UTR及284 bp的3′UTR,其中ORF编码一个具有135个氨基酸的蛋白质。BLAST分析结果显示漾濞大泡核桃JsPRP1编码的蛋白质与甜橙PRP (Citrus sinensis,XP_006477234.1)的氨基酸序列具有84%的相似性。JsPRP1含有N端信号肽、富含脯氨酸重复区域和C端8个特定排列的半胱氨酸,具有典型的PRPs特征,这表明其属于漾濞大泡核桃中的富含脯氨酸蛋白。超表达序列表SEQ ID NO :1所示序列可以增强烟草对胶孢炭疽菌(Colletotrichum gloeosporioides)、核盘菌(Sclerotinia sclerotiorum)、葡萄座腔菌(Botrosphaeria dothidea)以及串珠状赤霉菌(Gibberella moniliformis)的抗性。
上述JsPRP1基因可以应用于提高烟草的抗真菌特性,具体操作如下:
(1)采用扩增JsPRP1的特异引物,从接种胶孢炭疽菌后的漾濞大泡核桃叶中提取总RNA,通过逆转录-聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR)扩增出JsPRP1的全长编码区,然后将其连接到pMD-18T载体上,经测序获得具有目的基因的克隆。
(2)用限制性内切酶BamHI和EcoRI酶切pMD18-T-JsPRP1载体和植物表达载体pCAMBIA2300S,通过胶回收得到目的基因片段和载体大片段。再将所获得JsPRP1基因片段与pCAMBIA2300S载体片段连接,构建植物超表达载体。之后将所构建的重组载体通过根癌农杆菌介导转入烟草中表达。
(3)以重组载体T-DNA上具有的抗性标记筛选转化子,并通过PCR以及RT-PCR检测得到阳性转基因植株,分析转基因植株对于病原真菌的抗性,最后筛选出对真菌抗性明显增强的转基因植株。
本发明为提高植物对真菌病害的抗性提供了一种新的方法,通过基因工程手段培育抗病植物可以克服传统育种的不足,不仅育种周期缩短,而且操作简单,容易获得高抗材料。本发明中来自漾濞大泡核桃的JsPRP1基因能增强植物对几种病原真菌的抗性,将该基因导入烟草中,可以产生具有真菌抗性的新品种和新材料。利用基因工程技术培育抗性植物品种和材料具有明显的优势和不可取代的重要性。它不仅可以为大规模生产作物、花卉等提供方便,减少化学农药的使用,还可以为农业生产节约成本、减少环境污染,因此本发明具有广阔的市场应用前景。
附图说明
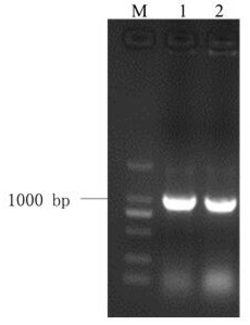
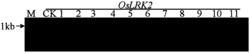
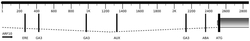
图1是本发明中部分JsPRP1转基因烟草基因组DNA的PCR检测结果,其中Marker:DL2000 DNA Marker (大连宝生物),由2,000 bp、1,000 bp、750 bp、500 bp、250 bp以及100 bp六条DNA片段组成;正对照:质粒pMD18-T-JsPRP1为模板的PCR反应;WT:非转基因烟草(野生型)总DNA为模板进行的PCR;
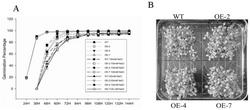
图2是本发明中部分阳性JsPRP1转基因烟草中JsPRP1转录水平的表达分析结果图,其中Marker:DL2000 DNA Marker(大连宝生物);WT:非转基因烟草总RNA逆转录cDNA为模板的PCR产物;正对照:质粒pMD18-T-JsPRP1为模板的PCR产物;
图3是本发明中JsPRP1转基因烟草体外抗真菌活性的抑菌效果图;其中a、b、c、d图示中的真菌分别是核盘菌、胶孢炭疽菌、串珠状赤霉菌以及葡萄座腔菌;WT为野生型烟草的总蛋白;CK为空白对照,即无蛋白对照(用于提取蛋白的缓冲液)。
具体实施方式
下面通过附图和实施例对本发明进一步说明,但本发明保护范围不局限于所述内容,本实施例中方法如无特殊说明的均按常规方法操作,所用试剂如无特殊说明的采用常规试剂或按常规方法配置的试剂。
实施例1:JsPRP1全长cDNA克隆以及序列分析
用胶孢炭疽菌接种漾濞大泡核桃,用接种后4 h的叶提取总RNA,用液氮将处理过的漾濞大泡核桃的叶研磨成粉末,然后转入离心管中,采用异硫氰酸胍法提取总RNA。采用逆转录酶M-MLV (promega)以总RNA为模板合成cDNA第一链,反应体系和操作过程为:取5 μg total RNA,依次加入50 ng oligo (dT),2 μL dNTP Mix (2.5 mM each),用DEPC水将反应体积补齐至14.5 μL;混匀后,70℃加热变性5 min后迅速在冰上冷却5 min,然后依次加入4 μL 5×First-stand buffer、0.5 μL RNasin (200U)、1 μL M-MLV (200U),混匀并简短离心,42℃温浴1.5 h,取出后70℃加热10 min,终止反应。cDNA第一链合成后置于-20℃保存备用。
以合成的第一链cDNA为模板,扩增目的基因JsPRP1,所用上下游引物序列分别为 -CAAGCAGTGGTATCAACGCAGAG- 及 -ACCTTGCATCCATCGAAACAAT- 。采用AdvantageTM 2 PCR Enzyme (Clontech)扩增出目的基因。PCR反应条件:95℃ 1 min;94℃ 30 s,60℃ 30 s,72℃ 45 s,32个循环;72℃ 5 min。反应体系(20 μL)为1 μL cDNA、2 μL 10×Advantage 2 PCR Buffer、1.8 μL dNTP Mix (10mM each)、0.2 μL 正向引物(10 μM)、0.2 μL 反向引物(10 μM)、0.2 μL Advantage 2 PCR Polymerase Mix、14.6 μL PCR-Grade water。PCR结束后,取8 μL进行琼脂糖凝胶电泳,用以检测扩增产物的特异性以及大小。
所得到PCR产物只有一条DNA带,故直接对PCR产物进行TA克隆,使用的试剂盒为pMD18-T vector kit (大连宝生物),反应体系和操作过程为:取1.5 μL PCR产物,依次加入1 μL pMD18-T vector (50 ng/μL)和2.5 μL 2×Ligation solution I,混匀后置于16℃过夜反应。通过热激转化法将连接产物转入大肠杆菌DH5α感受态中。用含有氨苄青霉素(ampicillin,Amp)的LB固体培养基筛选阳性克隆。挑选若干个单菌落,摇菌后用扩增JsPRP1的特异引物检测多克隆位点插入JsPRP1的克隆。将得到的阳性克隆进行测序,最终获得的JsPRP1全长cDNA为815 bp,通过NCBI ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析发现其包含一个408 bp的开放读码框(见序列表)。JsPRP1编码一个含135个氨基酸的蛋白质JsPRP1,其分子量约为14.17 KDa,等电点为8.91。借助生物信息学软件SignalP 4.1分析JsPRP1编码的蛋白序列,检测其是否具有N端信号肽。结果显示在JsPRP1的N端存在信号肽,因此推测该蛋白是分泌蛋白。
实施例2:植物超表达载体构建
采用SanPrep柱式质粒DNA小量抽提试剂盒(上海生工)提取插入JsPRP1的大肠杆菌质粒pMD18-T- JsPRP1以及植物表达载体pCAMBIA2300S质粒,取1 μL用于琼脂糖凝胶电泳以检测所提取质粒的完整性及浓度高低。用限制性内切酶EcoRI (TaKaRa)和BamHI (TaKaRa)分别对质粒pMD18-T-JsPRP1和pCAMBIA2300S进行双酶切(100 μL体系),反应体系和操作过程为:分别取20 μL pMD18-T-JsPRP1和pCAMBIA2300S质粒、依次加入10 μL 10×K buffer、5 μL EcoRI、5 μL BamHI、60 μL ddH2O,混匀后短时离心,置于37℃过夜反应。将所有酶切产物进行琼脂糖凝胶电泳,然后使用SanPrep柱式DNA胶回收试剂盒 (上海生工)对JsPRP1片段和pCAMBIA2300s载体大片段分别进行胶回收,取1 μL回收产物通过琼脂糖凝胶电泳检测回收片段的大小以及浓度,置于-20℃保存备用。
利用T4 DNA Ligase (TaKaRa),将回收的JsPRP1DNA片段和pCAMBIA2300S载体片段连接起来,反应体系(20 μL)和操作过程为:取10 μL JsPRP1DNA片段依次加入2 μL pCAMBIA2300S载体DNA、2 μL 10×T4 DNA Ligase Buffer、1 μL T4 DNA Ligase、5 μL ddH2O,混匀后短时离心,然后16℃水浴过夜反应。接着采用热激转化法将连接产物转入大肠杆菌DH5α中,用含有50 mg/L卡那霉素(kanamycin,Km)的固体培养基筛选阳性克隆。挑选单菌落摇菌,以菌液为模板用扩增JsPRP1的特异引物进行PCR,挑选出JsPRP1与pCAMBIA2300S成功连接的克隆,并向检测得到的阳性菌株中加入甘油并置于-80℃保存备用。
采用SanPrep柱式质粒抽提试剂盒(上海生工)提取并纯化上述大肠杆菌DH5α中的pCAMBIA2300S-JsPRP1质粒。随后用液氮冻融法将上述构建的植物表达载体pCAMBIA2300S-JsPRP1转入所制备的根癌农杆菌LBA4404感受态细胞中。操作步骤为:取2 μg pCAMBIA2300S-JsPRP1质粒加入含有200 μL感受态细胞的离心管中,轻轻混匀后冰浴5 min,随后转入液氮中冷冻1 min,然后迅速置于37℃水浴5 min,再冰浴2 min,之后加入500 μL LB液体培养基于28℃振荡培养4 h。将活化后的农杆菌涂于含有50 mg/L Km的LB固体培养基上,28℃倒置培养。挑选单菌落摇菌,再用扩增JsPRP1的特异性引物进行PCR反应,检测pCAMBIA2300S-JsPRP1是否转入农杆菌中。对于阳性克隆,加入甘油后置于-80℃保存备用。
实施例3:农杆菌介导的植物遗传转化以及转基因植物筛选
本实验的转基因受体是烟草(Nicotiana tabacum L.)。将烟草种子用75%的酒精浸泡30 s,无菌水洗涤后用0.1%的HgCl2浸泡8 min,然后再用无菌水洗涤若干次,播种于1/2 MS培养基上,28℃暗培养5-8 d,发芽后转至光照培养箱(25℃,16 h/d光照),以后每月用MS培养基继代一次。
从-80℃冰箱中取出保存的含有pCAMBIA2300S-JsPRP1质粒的农杆菌LBA4404菌种,取20 μL接种于5 mL含有50 mg/L Km和20 mg/L利福平的LB液体培养基中,28℃培养至培养基浑浊。吸取1 mL浑浊的菌液至含有50 mg/L Km的LB固体培养基上,28℃培养48 h。随后将LB固体培养基上的农杆菌刮下适量接种于附加有20 mg/L的乙酰丁香酮的MGL液体培养基中,28℃振荡培养5-8 h以活化农杆菌。
取烟草无菌烟草幼嫩叶片切成约1 cm2的叶盘,完全浸泡于上述含有活化农杆菌的MGL液体培养基中,25℃浸染15 min。用无菌滤纸吸干叶盘表面的菌液,将叶盘置于共培养基上,22℃无光条件下共培养2天。烟草转化的共培养基为MS+0.02 mg/L 6-BA+2.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂。
将共培养后的叶盘转到加有抗生素的MS筛选培养基中分化成苗,同时筛选转基因植株。烟草筛选培养基为MS+0.5 mg/L 6-BA+0.1 mg/L NAA+30 g/L蔗糖+6 g/L琼脂+50 mg/L Km+200 mg/L 头孢霉素(cefotaxime sodium salt,Cef);筛选培养时将培养瓶转移至光照培养箱培养(25℃,16 h/d光照,8 h/d黑暗)。待烟草长出芽后用含有50 mg/L Km和200 mg/L Cef的MS培养基继代培养。因烟草愈伤分化率较高,故需要对再生植株进行进一步筛选。将烟草再生苗移至含有50 mg/L Km的MS培养基上使其生根,最后选用生根较好的再生苗做进一步的检测。
采用CTAB法提取转基因烟草植株叶片的基因组DNA,取1 μL所得基因组DNA进行琼脂糖凝胶电泳检测其完整性和浓度。以转基因植株的基因组DNA为模板用JsPRP1的特异引物进行PCR反应。PCR结束后,取8 μL产物用于琼脂糖凝胶电泳以检测阳性转基因植株。部分烟草转基因植株的扩增结果如图1所示,JsPRP1转基因烟草共筛选到32株阳性转基因植株。
实施例4:转基因烟草中JsPRP1的表达分析以及转基因植株抗真菌活性分析
分别取阳性转基因植株以及非转基因烟草(野生型)的嫩叶提取总RNA,逆转录生成cDNA第一链,并以此为模板用扩增JsPRP1的特异引物进行PCR,根据PCR结果分析各转基因植株中JsPRP1转录水平的表达量。总RNA提取以及RT-PCR的方法与实施例1中相同。PCR结束之后,取8 μL用于琼脂糖凝胶电泳,部分单株的检测结果如图2所示,共检测到20个转基因单株中JsPRP1在转录水平大量表达,这些单株的编号为1~20。
将实验室保存的几种真菌接种于PDA固体培养基(200 g/L马铃薯,15 g/L琼脂,20 g/L葡萄糖)上,28℃暗培养,待菌落生长至直径约为2~3cm时添加蛋白,分析转基因植株体外抗真菌活性。胶孢炭疽菌(C. gloeosporioides) 、核盘菌(S. scleroterum)、葡萄座腔菌(B. dothidea)、和串珠状赤霉菌(G. moniliformis)。为了防止其它杂菌污染所提取的蛋白,整个植物蛋白提取过程均是无菌操作。首先取1 g转基因烟草单株(编号分别为3、4、6、10)及野生型叶片放入研钵中,加入1 mL蛋白提取液(1 M NaCl,0.1 M 乙酸钠,1% PVP,pH6.0),充分研磨。转入1.5 mL离心管中,混匀后4℃静置过夜。4℃离心30 min (12,000 g),取上清于新的1.5 mL离心管中,并取适量用紫外分光光度仪测定总蛋白浓度。将转基因和野生型植株的总蛋白浓度调整至0.2 μg/μL,然后分别取20 μL滴于各真菌培养基的无菌滤纸上。在每个真菌的平板上除了添加不同转基因烟草植株的总蛋白,同时平行添加野生型烟草的总蛋白和空白对照(蛋白提取液)。28℃培养几天后观察各处理真菌生长的情况,并据此来评价JsPRP1转基因烟草的体外抗真菌活性。结果如图3所示,JsPRP1转基因烟草蛋白对胶孢炭疽菌、核盘菌、葡萄座腔菌以及串珠状赤霉菌的生长具有明显的抑制作用。
序列表(SEQ ID)
<110> 昆明理工大学
<120> 漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1及应用
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 815
<212> DNA
<213> Juglans sigillataDode
<220>
<221> mRNA
<222> (1)..(815)
<220>
<221> 5'UTR
<222> (1)..(123)
<220>
<221> CDS
<222> (124)..(531)
<220>
<221> 3'UTR
<222> (532)..(815)
<400> 1
ctaatacgac tcactatagg gcaagcagtg gtatcaacgc agagtacatg gggactcaaa 60
tttgctctag aggcctttcc ctatttcaca aaccatacat taattatttt ggttttctgt 120
gaaatggctt ctaaggctct agcatctggt gccctcctcc tctctctcaa cctccttttc 180
tttgtgatgg tcagttccac ttatgtccct tgcccaccac ccccaaaggg tcacaaacac 240
caccatcaca agccctcccc ggcaccgaag cccacttgcc ccaaggacac ccttaagttg 300
ggggtgtgtg ccaacttgtt gaatgacttg gtacaccttg ttgtcggtac cccaccaaag 360
accccttgct gcagcctcat tgagggtctt gtggatctcg aagcagctgt ttgcctttgc 420
accgccatta aagctaatat cttgggcatc aaccttaacg tccccctgtc attgagcttg 480
ctgctcaact actgcggaaa gaaggtcccc tctggcttcc agtgcgcata aatctacact 540
agatttcacc atgtatcgtt tactatttat tttaatttcc cacttcccca ttgatccctt 600
taatcgtgtg attgggcttt ggcttgttta tttacgtatg ctataagcat ttatatatca 660
ataatttgtt gctgtttcat tgtttcgatg gatgcaaggt ttctcacctt catctatttt 720
caactaagta gggatcatct tatgctgtaa aaataaactg acactgtttc agaatgaagt 780
gaactaaaaa aaaaaaaaaa aaaaaaaaaa aaaaa 815
<210> 2
<211> 135
<212> PRT
<213> Juglans sigillataDode
<400> 2
Met Ala Ser Lys Ala Leu Ala Ser Gly Ala Leu Leu Leu Ser Leu Asn
1 5 10 15
Leu Leu Phe Phe Val Met Val Ser Ser Thr Tyr Val Pro Cys Pro Pro
20 25 30
Pro Pro Lys Gly His Lys His His His His Lys Pro Ser Pro Ala Pro
35 40 45
Lys Pro Thr Cys Pro Lys Asp Thr Leu Lys Leu Gly Val Cys Ala Asn
50 55 60
Leu Leu Asn Asp Leu Val His Leu Val Val Gly Thr Pro Pro Lys Thr
65 70 75 80
Pro Cys Cys Ser Leu Ile Glu Gly Leu Val Asp Leu Glu Ala Ala Val
85 90 95
Cys Leu Cys Thr Ala Ile Lys Ala Asn Ile Leu Gly Ile Asn Leu Asn
100 105 110
Val Pro Leu Ser Leu Ser Leu Leu Leu Asn Tyr Cys Gly Lys Lys Val
115 120 125
Pro Ser Gly Phe Gln Cys Ala
130 135
<210> 3
<211> 23
<212> DNA
<213> 人工序列
<400> 3
caagcagtgg tatcaacgca gag 23
<210> 4
<211> 22
<212> DNA
<213> 人工序列
<400> 4
accttgcatc catcgaaaca at 22
漾濞大泡核桃富含脯氨酸蛋白基因JsPRP1及应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0