





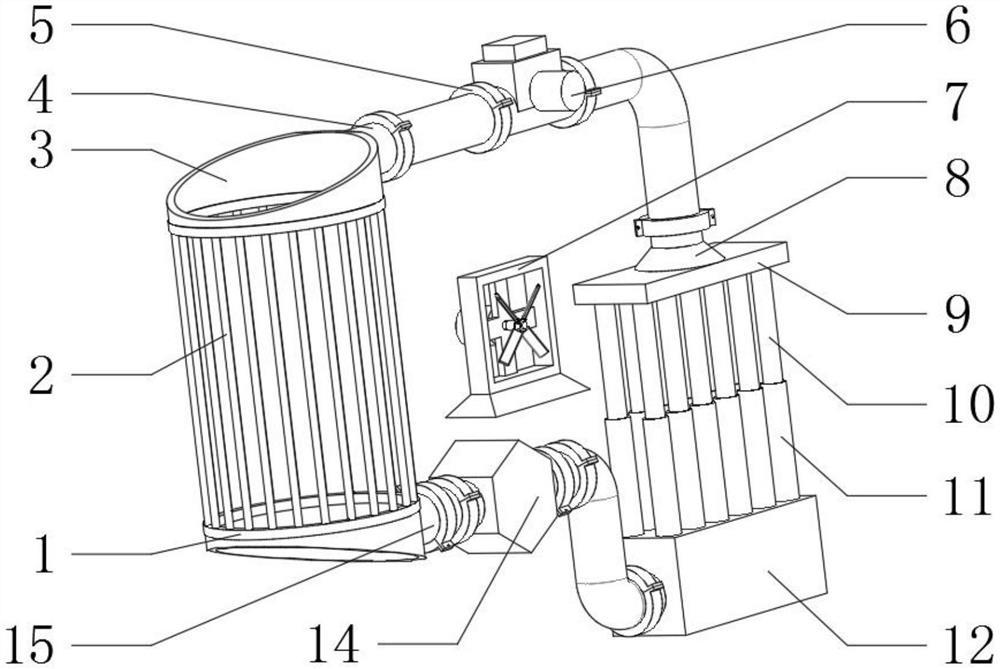





专利摘要
本文提供了用于在受试者组织损伤后促进伤口愈合和组织再生的组合物和方法。
权利要求
1.一种分离多肽,包含α连接蛋白的羧基端氨基酸序列或其保守变体,其中所述多肽不含全长的α连接蛋白。
2.权利要求1的多肽,其中所述多肽包含所述α连接蛋白的羧基端的4至30个连续氨基酸。
3.权利要求1的多肽,其中所述多肽包含:
(a)羧基端的II型PDZ结合基序;
(b)与PDZ结合基序毗邻的铰链基序,包含脯氨酸,甘氨酸,或脯氨酸和甘氨酸残基;以及
(c)与所述铰链残基毗邻的带正电荷残基(K、R、D、E)。
4.权利要求1的多肽,其中所述α连接蛋白选自连接蛋白39、连接蛋白40.1、连接蛋白47、连接蛋白46.6、连接蛋白30.2、连接蛋白31.9、连接蛋白44、连接蛋白33、连接蛋白36、连接蛋白35、连接蛋白37、连接蛋白39、连接蛋白39.9、连接蛋白40、鸡连接蛋白42、连接蛋白45.6、连接蛋白43、连接蛋白43.4、连接蛋白44.2、连接蛋白44.1、人连接蛋白45.6、连接蛋白46、连接蛋白56、连接蛋白49、连接蛋白50。
5.权利要求1的多肽,其中组织损伤后,所述多肽减轻炎症、促进愈合、减少瘢痕形成、增加抗拉强度并促进复杂组织再生。
6.权利要求1的多肽,其中所述多肽抑制连接蛋白复合蛋白与α连接蛋白的羧基端的结合,其中所述连接蛋白复合蛋白为ZO-1或肾胚细胞瘤过表达蛋白(NOV)。
7.权利要求4的多肽,其中所述α连接蛋白为Cx43。
8.权利要求7的多肽,其中所述多肽包含选自SEQ ID NO:1和SEQID NO:2的氨基酸序列。
9.权利要求2的多肽,其中所述多肽包含α连接蛋白羧基端的4至30个氨基酸内的保守氨基酸置换。
10.权利要求9的多肽,其中所述多肽包含与SEQ ID NO:1具有至少65%序列同一性的氨基酸序列。
11.权利要求9的多肽,其中所述多肽包含选自SEQ ID NO:3和SEQID NO:4的氨基酸序列。
12.权利要求4的多肽,其中所述α连接蛋白为Cx40。
13.权利要求12的多肽,其中所述多肽包含氨基酸序列SEQ ID NO:5。
14.权利要求1的多肽,还包含细胞内化序列。
15.权利要求14的多肽,其中所述细胞内化序列包含选自下列蛋白质的氨基酸序列:触角足蛋白、TAT、HIV-Tat、穿膜肽、Antp-3A(Antp突变体)、Buforin II、Transportan、MAP(模式两亲肽)、K-FGF、Ku70、朊病毒、pVEC、Pep-1、SynB1、Pep-7、HN-1、BGSC(双-胍-亚精胺-胆固醇)和BGTC(双-胍-Tren-胆固醇)。
16.权利要求15的多肽,其中所述氨基酸序列来自触角足蛋白,并包含氨基酸序列SEQ ID NO:7。
17.权利要求16的多肽,包含选自SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11和SEQ ID NO:12的氨基酸序列。
18.一种编码权利要求1的多肽的分离核酸。
19.权利要求18的分离核酸,其中所编码的多肽包含选自SEQ ID NO:1、SEQ D NO:2、SEQ ID NO:3、SEQ ID NO:4和SEQ ID NO:5的氨基酸序列。
20.权利要求19的分离核酸,包含核酸序列SEQ ED NO:6。
21.一种编码权利要求15的多肽的分离核酸。
22.权利要求21的分离核酸,其中所编码的多肽包含选自SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11和SEQ ID NO:12的氨基酸序列。
23.权利要求22的分离核酸,包含核酸序列SEQ ID NO:13。
24.权利要求19或22的分离核酸,其中所述核酸与表达调控序列有效连接。
25.一种载体,包含与表达调控序列有效连接的权利要求19或22的核酸。
26.权利要求25的载体,其中所述载体为病毒。
27.一种包含权利要求19或22的核酸的细胞。
28.一种包含权利要求25的载体的细胞。
29.一种包含权利要求19或22的核酸的生物。
30.一种包含权利要求25的载体的生物。
31.一种组合物,包含存在于可药用载体中的权利要求1或14的多肽。
32.一种组合物,包含存在于可药用载体中的权利要求19或22的核酸。
33.一种促进受试者组织损伤后的愈合的方法,包括将权利要求31或32的组合物给予所述受试者。
34.一种减少受试者组织损伤后的瘢痕组织形成的方法,包括将权利要求31或32的组合物给予所述受试者。
35.一种促进受试者组织损伤后的组织再生的方法,包括将权利要求31或32的组合物给予所述受试者。
36.一种增加受试者组织损伤后的抗拉强度的方法,包括将权利要求31或32的组合物给予所述受试者。
37.一种增强受试者组织损伤后的干细胞分化的方法,包括将权利要求31或32的组合物给予所述受试者。
38.一种减轻受试者的炎症的方法,包括将权利要求31或32的组合物给予所述受试者。
39.一种减少受试者纤维化组织形成的方法,包括将权利要求31或32的组合物给予所述受试者。
40.一种抑制受试者体内转化细胞的增殖的方法,包括将权利要求31或32的组合物给予所述受试者。
41.一种抑制受试者体内转化细胞的转移性迁移的方法,包括将权利要求31或32的组合物给予所述受试者。
42.权利要求33-37任一项的方法,其中所述组织损伤是由于下列因素所致:刮伤,切伤,裂伤,挤压伤,加压伤,牵张性损伤,咬伤,擦伤,枪弹伤,爆炸伤,身体穿孔,刺伤,烧伤,风吹性皮肤伤,晒伤,化学烧伤,外科伤口,外科手术,医学介入,细胞、组织或器官移植后的宿主排斥现象,药物作用,药物副作用,褥疮,放射性损伤,化妆品引起的皮肤损伤,内脏损伤,疾病过程(如哮喘、癌症),感染,传染源,发育过程,化脓过程(如痤疮),遗传异常,发育异常,环境毒素,变应原,头皮损伤,面部损伤,颚损伤,脚损伤,脚趾损伤,手指损伤,骨损伤,性器官损伤,关节损伤,排泄器官损伤,眼损伤,角膜损伤,肌肉损伤,脂肪组织损伤,肺损伤,气道损伤,疝气,肛门损伤,痔疮,耳损伤,视网膜损伤,皮肤损伤,腹部损伤,手臂损伤,腿损伤,运动损伤,背部损伤,产伤,早产损伤,毒性咬伤,螫伤,腱损伤,韧带损伤,心脏损伤,心瓣膜损伤,血管系统损伤,软骨损伤,淋巴系统损伤,颅脑创伤,脱臼,食管穿孔,瘘,指甲损伤,异物,骨折,冻伤,手伤,热应激损伤,撕裂,颈损伤,自身致残,休克,创伤性软组织损伤,脊髓损伤,脊柱损伤,扭伤,劳损,腱损伤,韧带损伤,软骨损伤,胸部损伤,牙损伤,创伤,神经系统损伤,衰老,动脉瘤,中风,消化道损伤,梗塞和缺血性损伤。
说明书
技术领域促进伤口愈合和组织再生的组合物和方法
技术背景背景技术
普通儿童都知道若石龙子蜥蜴失去尾巴,那么它最终会再长出一条尾巴。另外,习惯于研究这类事物的儿童和成年人都非常清楚,很多低等动物在损伤之后能再生包括整个肢体和器官在内的非常复杂的结构。例如,鱼在相当一部分原来的心脏被切除之后,还能长成一个心脏(Poss et al.,2002)。在认识到心脏对于大部分动物每一分钟的生存都非常重要的情况下,这个结果确实出人意料。
但是,人受损伤后组织、肢体和器官的再生并不如在鱼中那么简单。尽管由机械损伤、疾病过程和其它原因所损害的人组织能愈合,但是对于复杂的组织结构和功能,即使能恢复,也很少能完全恢复。不同地,人和其它高等脊椎动物几乎所有的受损伤组织的康复都受控于瘢痕组织的形成。其最常见的实例就是皮肤切伤或擦伤愈合后依然存在的变色的和纤维变性的瘢痕。还未得到充分地认识是,脑或脊髓损伤后神经胶质瘢痕组织的形成是中枢神经系统损害后神经功能恢复的主要障碍之一(Silver andMiller JH,2004)。目前还没有在损伤后治疗或预防这种瘢痕形成以及促进复杂组织结构和功能的再生的方法。
发明内容发明内容
提供了一种包含α连接蛋白的羧基端氨基酸序列(在本文中还被称为α连接蛋白羧基端(ACT)多肽)或其保守变体的分离多肽。
本文提供了一种促进受试者组织损伤后的伤口愈合的方法,包括将本文所提供的一种或多种存在于可药用载体中的组合物(如多肽、核酸或载体)给予受试者。
所公开的方法和组合物的其它优点一部分将在随后的描述中进行阐述,一部分可从描述中得到理解或者通过实施所公开的方法和组合物获知。所公开方法和组合物的优点可通过在所附权利要求书中特别指出的要素和组合实现和获得。应理解的是,上面的概述以及随后的详细描述仅仅是示例性和解释性的,无意于对所要求保护的发明进行限制。
附图说明附图说明
并入本说明书并构成本说明书一部分的附图与说明书一起对所公开方法和组合物的几个实施方案进行了阐述,用来解释所公开方法和组合物的原理。
图1示出α连接蛋白羧基端(ACT)多肽提高了培养的新生肌细胞中Cx43间隙连接形成的程度。根据标准方法培养新生大鼠心脏的肌细胞,直至在组织培养皿上形成接近汇合的单层。随后将培养物在含有如下成分的培养基中再培养五天:(a)30μM ACT 1肽(SEQ ID NO:2)、(b)30μM非活性对照肽(SEQ ID NO:55)或(c)不含ACT肽的或对照的磷酸盐缓冲盐水(PBS)。实验期间每24小时更换添加了肽或载体对照的培养基。(a)表明相对于对照条件(b)和(c),ACT肽大大提高了肌细胞之间Cx43间隙连接形成(箭头示出的点和线)的程度。许多表达Cx43的细胞类型也在对ACT应答时增加其Cx43间隙连接形成。
图2示出ACT肽抑制划伤的转化成纤维细胞(NIH-3T3细胞)的增殖和迁移。用ACT 1肽(SEQ ID NO:2)将NIH-3T3单层预处理24小时,并用p200移液器吸头将其“划伤”。随后在(a,b)30μM ACT 1肽(SEQ ID NO:2)、(c,d)30μM非活性对照肽(SEQ ID NO:55)或(e,f)不含ACT肽或对照肽的载体对照溶液的存在下使“划伤”愈合24小时。ACT肽处理的细胞的“划伤”在24小时后保持相对未愈合状态(a),鲜有细胞(大箭头)在最初“划伤”边缘内的区域(即小的黑色箭头标记的区域内)重新长入。相反,在(c)和(e)的对照条件中,大量细胞(大箭头)已在最初的“划伤”区域内重新长入。“划伤”区域细胞的重新长入一部分是通过逐渐行至“划伤”区域的转化细胞的迁移。附图(b)、(d)和(f)示出“划伤”区域或损伤边缘的细胞的增殖细胞核抗原(PCNA)免疫标记。ACT肽处理的细胞(b)仅表现出与本底和未增殖情况相一致的较低亮度。仅在(d)和(f)所示的两种对照条件下才观察到显著标记的增殖细胞(白色箭头)。这表明ACT肽也抑制了转化细胞的增殖。
图3示出在实验细胞模型中对损伤后ACT肽抑制迁移的定量结果。NIH-3T3成纤维细胞被“划伤”并将其置于图2所述的30μM ACT 1肽(SEQID NO:2)或对照条件中达24小时。图a示出24小时后ACT肽处理的细胞和非活性肽处理的对照细胞的损伤边缘。该细胞用荧光鬼笔环肽标记以助于观察。ACT肽处理的细胞显示出划伤区域(白色双头箭头)的细胞重新长入水平较低。图b示出24小时后重新长入划伤区域的细胞的面积百分比柱状图。ACT肽存在时损伤区域细胞的减少是显著的,p值小于0.000001。
图4示出在实验细胞模型的上皮细胞WB-F344中,有效连接启动子的编码ACT肽的多核苷酸的表达可抑制划伤后的迁移。WB-F344细胞是通过用致癌剂处理分离的大鼠肝细胞所得到的转化大鼠上皮细胞系(Tsao et al.,1984;Hayashi et al.,1997;Hayashi et al.,1998;Hayashi et al,2001)。用cDNA表达质粒构建体转染WB-F344细胞,并使用标准方法在抗生素条件下对其进行筛选,以形成这样一种细胞系,所述细胞系可稳定表达有效连接启动子序列的编码ACT肽的多核苷酸(SEQ ID NO.6),或者可稳定表达作为对照的有效连接启动子序列的绿色荧光蛋白(GFP)多核苷酸。编码ACT肽的多核苷酸也编码GFP。因此,ACT肽的表达可在光学显微镜下通过标准GFP荧光光学法进行分析。(a)和(b)示出表达GFP和羧基端ACT肽序列(a)或仅表达GFP(b)的WB-F344细胞系的GFP荧光高倍放大图。将WB-F344细胞系接近汇合的细胞单层“划伤”并使其“愈合”24小时。与用载体或非活性对照肽处理的NIH-3T3细胞的对照情况相似,表达GFP的对照上皮细胞系重新长入划伤区(d)。但是,在稳定表达有效连接启动子序列的编码ACT肽的多核苷酸的上皮细胞系中,划伤细胞的重新长入受到抑制(c)。
图5示出ACT肽在新生小鼠切开性皮肤损伤后减轻炎症、促进愈合并减少瘢痕形成。通过低体温使新生小鼠脱敏。使用手术刀穿过肩胛骨之间的背中线的整层皮肤(下至下层肌肉水平)制成一个4mm长的切开性皮肤损伤。然后将30μl不含(对照)或含有溶解的ACT 1肽(SEQ ID NO:2)(浓度为60μM)的20%Pluronic(F-127)凝胶溶液施用至切口损伤处。随后,对照或含有ACT肽的凝胶在第一次施用后24小时再次施用。第二次施用之后不再施用对照和含ACT肽的凝胶。到48小时时,与没有接受ACT肽的对照损伤(b)相比,ACT肽处理的损伤(a)显著地闭合更好、发炎较少、肿胀较轻(注意伤口边缘的隆起),并且通常愈合形态更好。对照和ACT肽和对照处理的伤口之间炎症、肿胀和愈合的差异在72小时(c、d)和96小时(e、f)时保持一致。第7天时,ACT肽伤口(g)的形态较对照肽处理的损伤(h)的要平滑并且瘢痕较少。注:示出的是同一动物的同一损伤在愈合期间的不同时间点的图。
图6示出ACT肽在成年小鼠较大切除性皮肤损伤后减轻炎症、促进愈合并减少瘢痕形成。使用精细的手术剪在麻醉的成年小鼠肩胛骨之间的背中线(下至皮肤下层的肌肉)制成8mm宽的环形切除性皮肤损伤(即(a)和(b)所示)。通过在塑料片上剪出8mm宽环形模板标记出损伤边界。然后将100μl不含(对照)或含有溶解ACT 1肽(SEQ ID NO:2)(浓度为100μM)的30%Pluronic凝胶溶液施用至切除性损伤处。随后,对照或含有ACT肽的凝胶在第一次施用后24小时再次施用。第二次施用之后不再施用对照和含ACT肽的凝胶。在14天期间,ACT肽处理的较大切除性损伤(a,c,e,g,i)较没有ACT肽处理的对照损伤(b,d,f,h,j)闭合更快、外在炎症减轻、愈合更快并且瘢痕较少。事实上,第14天的对照损伤依然显示出有部分结痂,这表明损伤的快速愈合并不彻底(j)。注:示出的是同一动物的同一损伤在愈合期间的不同时间点的图。
图7示出ACT肽在成年小鼠切除性皮肤损伤后减少炎症细胞的密度。在图6所述实验中的切除性损伤后24小时对其中一些小鼠的整个伤口部位进行皮肤活组织检查。图(a)和(b)分别为示出对照和ACT肽处理损伤的伤口中心附近的横截面的低倍放大视图。在两种情况下均可观察到由正常组织学形态的皮肤形成边界的伤口边缘(由小箭头标记)。黑色方框置于(a)和(b)图中的左侧伤口边缘上。这两个方框内的对照和ACT肽处理组织的组织学结构分别在(c)和(d)以较高放大倍数示出。最引人注意的是从损伤的基底部分向损伤的伤口边缘和损伤外表面投射的整齐排布的纤维物质(箭头所示)的“领状”组织。对照损伤(d)中整齐排布的纤维基质的形态较ACT肽处理损伤(c)的形态构成有序得多。此外,还有相当低密度的炎症细胞散布于ACT肽处理组织的纤维基质中。这在(e)和(f)得到确认,其中(d)和(c)示出的黑色方框内的组织切片区域分别以较高放大倍数示出。散布于整齐排列的纤维基质的炎症细胞包括肥大细胞、嗜中性粒细胞和巨噬细胞。对照损伤中的这些炎症细胞的密度较ACT肽处理损伤中的这些炎症细胞的密度高的多。
图8示出ACT肽在成年小鼠切除性皮肤损伤后促进愈合、减少瘢痕形成和促进复杂的组织结构再生。在图6所述实验的第14天结束时,对整个切除性损伤进行皮肤活组织检查并对这些皮肤样本的组织切片进行H&E组织化学染色。图(a)和(b)分别示出ACT肽和对照的伤口中心附近的横截面的低倍放大视图。在两种情况下均可观察到由正常组织学形态的皮肤形成边界的伤口边缘(由小箭头标记)。黑色方框置于(a)和(b)图中的每个损伤的中心附近。这两个方框内的ACT肽和对照组织的组织学结构分别在(c)和(d)以较高放大倍数示出。很显然,ACT肽处理损伤位置内的组织更为复杂。在ACT肽处理的伤口外表面有一层连续的上皮细胞,表明损伤表面的上皮再形成是完全的,纵使伤口中心附近的上皮组织还相对较薄(c)。已观察到再生毛囊是从包覆愈合损伤的新上皮组织中的干细胞重新分化的(c,小箭头)。相比较而言,对照损伤的损伤表面的上皮再形成是不完全的,并且没有毛囊在上皮组织中再生的迹象(d)。在ACT肽处理的损伤皮肤重新形成的上皮组织下层,观察到正常组织复杂性的相当程度的恢复,并且腺体结构、纤维组织和结缔组织、血管组织、肌肉和脂肪细胞都很明显(a,c)。如同毛囊一样,该组织复杂性是通过干细胞的分化再生的。相反,在对照损伤中,伤口组织完全由均一和大块的纤维化瘢痕组织(b,d)占据,同时,在该瘢痕组织内,组织结构的其它复杂性不太明显。
图9示出ACT肽在成年小鼠切除性皮肤损伤后减轻炎症、促进愈合和减少瘢痕形成。使用精细的手术剪在麻醉的成年小鼠颈部和(上侧)背部剪出两个小的(直径为5mm)切除性皮肤伤口。通过在塑料片上剪出5mm宽环形模板标记出损伤边界。然后将50-60μl不含(对照)或含有溶解的一种ACT肽(ACT 2-SEQ ID NO:1、ACT 1-SEQ ID NO:2、ACT 3-SEQ ID NO:3、ACT 4-SEQ ID NO:4或ACT 5-SEQ ID NO:5)(浓度为100μM)的20%Pluronic凝胶溶液施用至切除性损伤处。随后,对照或含有ACT肽的凝胶在第一次施用后24小时再次施用。第二次施用之后不再施用对照和含ACT肽的凝胶。可注意到,在240小时期间(10天),ACT 1(e-h)、ACT 2(i-l)、ACT 3(m-p)和ACT 5(u-x)肽处理的切除性损伤较没有ACT肽处理的对照损伤(a-d)闭合更快,外在炎症显著减轻,愈合更快并且瘢痕较少。在此期间,ACT4肽(q-t)相对于对照在愈合方面也表现出中等程度的改进,但是不如其它肽。注:示出的是同一动物的同一伤口在愈合期间不同时间点的图。
图10示出ACT肽在成年大鼠脑贯通伤(penetration injury)后减少形成神经胶质瘢痕的星形细胞的数量和密度。(b)和(c)为低倍放大视图,示出填充有ACT肽(100μM)加载体凝胶(b)或作为对照的胶原载体凝胶(c)的中空纤维膜(HFM)植入物周围的脑组织(皮质)切片。在对照组织中(c),在HFM所致损伤部位附近观察到高密度的免疫标记的GFAP阳性星形细胞。这些细胞的密度似乎在损伤远端略有下降。相反,在填充有ACT肽的HFM附近观察到密度低得多的GFAP阳性星形细胞(b)。事实上,GFAP阳性细胞的水平与正常未损伤脑组织中观察到的基本相同。图(b)和(c)白色方框内的组织区域分别在(d)和(e)中以高倍放大形式示出。在ACT肽处理的脑损伤中(d),可观察到GFAP阳性星形细胞与在对照损伤(e)中观察到的相比不仅较少,而且更小。
图11示出ATC肽在成年大鼠脑贯通伤后有助于神经元的维持以及神经元的再生。(a)和(b)为低倍放大视图,示出在脑贯通伤后一周填充有对照胶原载体凝胶或ACT肽加载体凝胶的HFM植入物(植入物或损伤边缘由箭头示出)周围的脑组织(皮质)切片。在对照组织(b)中,在HFM所致损伤部位附近观察到高密度免疫标记的GFAP阳性星形细胞和低密度NeuN免疫标记的神经元。这些细胞的密度似乎分别在损伤远端降低和升高。相反,在填充有ACT肽(a)的HFM附近(以及远端)观察到密度低得多的GFAP阳性星形细胞和更多的NeuN免疫标记神经元。(a)和(b)中HFM附近区域分别在(c)和(b)的高倍放大视图中示出。与ACT肽处理的组织(c)相比,在对照组织(d)中也观察到密度显著增加的GFAP阳性星形细胞和密度减少的NeuN阳性神经元。在含ACT肽的HFM附近观察到了补偿模式,NeuN阳性神经元明显多于星形细胞(c)。值得注意的是,(c)所示高倍放大视图表明,相对于对照(d),频繁出现处于细胞核分裂过程中的神经元。
具体实施方式具体实施方式
通过参照下文对具体实施方案和其中包含的实施例的详细描述并参照附图以及之前之后对其的描述,使所公开方法和组合物更容易理解。
提供了一种分离多肽,包含α连接蛋白的羧基端氨基酸序列(在本文中也成为α连接蛋白羧基端(ACT)多肽)或其保守变体。一方面,在组织损伤后,所提供的ACT多肽可减轻炎症、促进愈合、减少瘢痕形成、增加抗拉强度并促进复杂组织的再生。另一方面,所提供的多肽增大了由连接蛋白所形成的间隙连接通道聚集体的范围。
应理解的是,除非另外指出,所公开的组合物和方法并不限于特定的合成方法、特定分析技术或者具体试剂,因此可有所变化。还应理解的是,本文所使用的术语目的仅在于对具体的实施方案进行描述,而无意于对其进行限制。
公开了可用于所公开方法和组合物的、可与所公开方法和组合物一起使用的、可用于制备所公开方法和组合物的产品的和作为所公开方法和组合物的产品的物质、组合物和组分。在本文中公开了这些物质和其他物质,应当理解的是,在公开这些物质的组合、子集、相互作用和群组等的时候,尽管关于这些化合物的每种不同的个体和集体的组合和排列可能并未明确地公开,但每种都在此被特别地考虑和描述。例如,如果公开和讨论了一种载体,并且讨论了多种包括启动子在内的载体组分,则除非另外特别指明,启动子和其它载体组分的每一种组合和排列以及可能的改变均得到特别地考虑。因此,如果公开了一类分子A、B和C,并且还公开了一类分子D、E和F以及组合分子A-D的实例,则即使没有每个逐一列举,每个也单独和共同得到考虑。因此,在这个实例中,A-E,A-F,B-D,B-E,B-F,C-D,C-E以及C-F的每个组合也被特别地予以考虑,并且认为上述每一个组合也因为A、B和C;D、E和F;以及组合分子A-D的公开而被公开。同样,这些分子的任何子集或组合也被特别地予以考虑和公开。因此,例如,A-E,B-F和C-E的子群也被特别地予以考虑,并且因为A、B和C;D、E和F;以及组合分子A-D的公开而被公开。将这一概念适用于本申请的所有方面,包括但不限于制备和应用公开组合物的方法中的步骤。因此,如果有多个其他步骤可以实施,应当理解的是,所述其他步骤的每一步都可以以所公开方法的任何具体实施方案或实施方案的组合来实施,并且每种这样的组合都被特别地予以考虑和公开。
本文提供了多个序列,这些序列和其它序列可在Genbank(www.pubmed.gov)找到。本领域的技术人员知晓如何分辨序列偏差和差异以及如何将与组合物和方法相关的具体序列调整为其它相关序列。根据本文所公开以及本领域已知的信息,可对任何给定序列设计引物和/或探针。
本文所提供的多肽可为包含α连接蛋白羧基最末端多个氨基酸的任何多肽,其中所述多肽不包含全长α连接蛋白。因此,一方面,所提供的多肽不含α连接蛋白的胞质N端结构域。另一方面,所提供的多肽不含α连接蛋白的两个胞外结构域。另一方面,所提供的多肽不含α连接蛋白的四个跨膜结构域。另一方面,所提供的多肽不含α连接蛋白的胞质环状结构域。另一方面,所提供的多肽不含邻近第四跨膜结构域的α连接蛋白的胞质羧基端结构域的部分序列。α连接蛋白中存在保守脯氨酸或甘氨酸残基,它始终位于距离羧基最末端的氨基酸大约17至30个氨基酸的位置(表2)。例如,对于人Cx43,第363位氨基酸脯氨酸残基距离羧基最末端的异亮氨酸19个氨基酸。在另一个实例中,对于鸡Cx43,第362位氨基酸脯氨酸残基距离羧基最末端的异亮氨酸18个氨基酸。在另一个实例中,对于人Cx45,第377位氨基酸甘氨酸残基距离羧基最末端的异亮氨酸19个氨基酸。在另一个实例中,对于大鼠Cx33,第258位氨基酸脯氨酸残基距离羧基最末端的甲硫氨酸28个氨基酸。因此,另一方面,所提供的多肽不包含与α连接蛋白的所述保守脯氨酸或甘氨酸残基相邻的氨基酸。因此,所提供的多肽可包含α连接蛋白的羧基最末端的4至30个氨基酸,包括α连接蛋白的羧基最末端的4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30个氨基酸。
所提供的肽中α连接蛋白羧基最末端的氨基酸的侧翼可含有非α连接蛋白或非ACT肽连接蛋白氨基酸。本文提供了非α连接蛋白和非ACT连接蛋白氨基酸侧翼的实例。非ACT连接蛋白氨基酸的一个实例为人Cx43羧基端的20至120个氨基酸(SEQ ID NO:72)。另一个实例为鸡Cx43羧基端的20至120个氨基酸(SEQ ID NO:73)。另一个实例为人Cx45羧基端的20至120个氨基酸(SEQ ID NO:74)。另一个实例为鸡Cx45羧基端的20至120个氨基酸(SEQ ID NO:75)。另一个实例为人Cx37羧基端的20至120个氨基酸(SEQ ID NO:76)。另一个实例为大鼠Cx33羧基端的20至120个氨基酸(SEQ ID NO:77)。
非α连接蛋白的一个实例为增强型绿色荧光蛋白的239氨基酸序列(图4示出ACT1与GFP功能性融合;SEQ ID NO:78)。另一方面,由于已表明ACT1与GFP的239氨基酸序列的羧基端融合时具有功能(如图4),ACT肽的侧翼最多有至少239个氨基酸的非连接蛋白多肽时,ACT肽有望保持其功能。事实上,只要维持ACT肽序列作为某一给定多肽的游离羧基端,则所述ACT肽就能接近其靶标。因此,除所述ACT肽之外的氨基酸超过239个的多肽具有在损伤后减轻炎症、促进愈合、增加抗拉强度、减少瘢痕形成并促进复杂组织再生的功能。
连接蛋白是负责细胞间通讯的间隙连接通道的亚基蛋白(Goodenoughand Paul,2003)。基于核苷酸序列的保守模式,编码连接蛋白的基因可分为被称为α和β连接蛋白基因的两个家族。α连接蛋白羧基最末端的氨基酸序列的特征在于具有多个独特和保守的特征(参见表2)。这种构成上的保守与ACT肽的如下能力相一致:形成独特的3D结构、与多个伴侣蛋白相互作用、介导与脂质和膜的相互作用、与包括DNA在内的核酸相互作用、通过和/或阻断膜通道以及提供蛋白酶剪切、蛋白质交联、ADP-核糖基化、糖基化和磷酸化的共有基序。因此,所提供的多肽可与蛋白质的这样一种结构域相互作用,所述结构域通常介导所述蛋白与α连接蛋白的羧基端结合。例如,肾胚细胞瘤过表达蛋白(NOV)与Cx43羧基端结构域相互作用(Fu et al,J Biol Chem.2004 279(35):36943-50)。认为这种蛋白质和其它蛋白质可与α连接蛋白的羧基端相互作用,并且还可与其它蛋白质相互作用,形成大分子复合物。因此,所提供的多肽可抑制分子机制例如一种参与调节Cx43间隙连接通道的聚集的分子机制发挥作用。
本文所使用的“抑制”、“阻止”和“阻滞”意味着降低(减轻)活性、反应性,病症、疾病或其它生物参数。这可包括但不限于活性、反应性、病症或疾病的完全丧失或根除。这还可包括与天然或对照水平相比,活性、反应性、病症或疾病降低或了10%。因此,与天然或对照水平相比,该降低或减轻可为10、20、30、40、50、60、70、80、90、100%,或者这之间任何量的降低或缓解。
所提供多肽的ACT序列可来自任何α连接蛋白。因此,所提供多肽的α连接蛋白组分可来自人、鼠类、牛、单孔类(monotrene)、有袋类、灵长类、啮齿类、鲸类、哺乳动物、禽类、爬行类、两栖类、鱼类、脊索动物、原索动物或其它来源的α连接蛋白。
因此,所提供多肽可包含选自下列的连接蛋白的ACT:小鼠连接蛋白47、人连接蛋白47、人连接蛋白46.6、牛连接蛋白46.6、小鼠连接蛋白30.2、大鼠连接蛋白30.2、人连接蛋白31.9、狗连接蛋白31.9、绵羊连接蛋白44、牛连接蛋白44、大鼠连接蛋白33、小鼠连接蛋白33、人连接蛋白36、小鼠连接蛋白36、大鼠连接蛋白36、狗连接蛋白36、鸡连接蛋白36、斑马鱼连接蛋白36、石鮨连接蛋白35、石鮨连接蛋白35、蝾螈连接蛋白35、鲀属(Tetraodon)连接蛋白36、人连接蛋白37、黑猩猩连接蛋白37、狗连接蛋白37、仓鼠属(Cricetulus)连接蛋白37、小鼠连接蛋白37、黄金仓鼠属(Mesocricetus)连接蛋白37、大鼠连接蛋白37、小鼠连接蛋白39、大鼠连接蛋白39、人连接蛋白40.1、非洲蟾蜍属连接蛋白38、斑马鱼连接蛋白39.9、人连接蛋白40、黑猩猩连接蛋白40、狗连接蛋白40、牛连接蛋白40、小鼠连接蛋白40、大鼠连接蛋白40、仓鼠属连接蛋白40、鸡连接蛋白40、人连接蛋白43、长尾猴属(Cercopithecus)连接蛋白43、真兔属(Oryctolagus)连接蛋白43、掘地小粟鼠属(Spermophilus)连接蛋白43、仓鼠属连接蛋白43、毛跖鼠属(Phodopus)连接蛋白43、大鼠连接蛋白43、猪属(Sus)连接蛋白43、黄金仓鼠属连接蛋白43、小鼠连接蛋白43、豚鼠属(Cavia)连接蛋白43、牛连接蛋白43、猬属(Erinaceus)连接蛋白43、鸡连接蛋白43、非洲蟾蜍属连接蛋白43、真兔属连接蛋白43、鲤属(Cyprinus)连接蛋白43、斑马鱼连接蛋白43、红蚌鱼丹(Danio aequipinnatus)连接蛋白43、斑马鱼连接蛋白43.4、斑马鱼连接蛋白44.2、斑马鱼连接蛋白44.1、人连接蛋白45、黑猩猩连接蛋白45、狗连接蛋白45、小鼠连接蛋白45、牛连接蛋白45、大鼠连接蛋白45、鸡连接蛋白45、鲀属连接蛋白45、鸡连接蛋白45、人连接蛋白46、黑猩猩连接蛋白46、小鼠连接蛋白46、狗连接蛋白46、大鼠连接蛋白46、黄金仓鼠属连接蛋白46、仓鼠属连接蛋白46、鸡连接蛋白56、斑马鱼连接蛋白39.9、牛连接蛋白49、人连接蛋白50、黑猩猩连接蛋白50、大鼠连接蛋白50、小鼠连接蛋白50、狗连接蛋白50、绵羊连接蛋白49、黄金仓鼠属连接蛋白50、仓鼠属连接蛋白50、鸡连接蛋白50、人连接蛋白59或其它α连接蛋白。α连接蛋白的氨基酸序列为本领域所知并且包括表1中由编号所表示的氨基酸序列。
表1:α连接蛋白
因此,所提供多肽可包含氨基酸序列SEQ ID NO:1、SEQ ID NO:29、SEQ ID NO:30、SEQ ID NO:31、SEQ ID NO:32、SEQ ID NO:33、SEQ ID NO:34、SEQ ID NO:35、SEQ ID NO:36、SEQ ID NO:37、SEQ ID NO:38、SEQ ID NO:39、SEQ ID NO:40、SEQ ID NO:41、SEQ ID NO:43、SEQ ID NO:90或ID NO:91或它们的保守变体或片段。
α连接蛋白羧基最末端20-30个氨基酸的序列的特征在于独特和保守的构成。该独特和保守的构成包括II型PDZ结合基序(Φ-x-Φ;其中x=任何氨基酸,Φ=疏水性氨基酸;如表2黑体所示)以及与该基序相邻的脯氨酸(P)和/或甘氨酸(G)铰链残基,高频率出现的磷酸-丝氨酸(S)和/或磷酸-苏氨酸(T)残基,以及高频率出现的带正电的精氨酸(R)、赖氨酸(K)和带负电的天冬氨酸(D)或谷氨酸(E)氨基酸。对于很多α连接蛋白,P和G残基出现在与羧基端II型PDZ结合基序相邻的成簇基序(clustered motif)中(如表2斜体表示)。大部分α连接蛋白的S和T磷酸氨基酸通常还构成了成簇的重复样基序(如表2下划线所示)。具体到Cx43的情况时,该构成中羧基最末端20个氨基酸的90%由后七种氨基酸构成。在高度保守的序列的另一实例中,Cx43的ACT肽构成在人和鱼类之间高度保守(如比较表2中的人和斑马鱼的Cx43ACT序列)。在另一实例中,Cx45的ACT肽构成在人和鸟类之间高度保守(如比较表2中的人和鸡的Cx45ACT序列)。在另一实例中,Cx36的ACT肽构成在灵长类和鱼类之间高度保守(如比较表2中的黑猩猩和斑马鱼的Cx36ACT序列)。
表2.α连接蛋白羧基端(ACT)氨基酸序列
因此,一方面,所提供的多肽包含选自下列的一个、两个、三个或全部氨基酸基序:1)II型PDZ结合基序;2)脯氨酸(P)和/或甘氨酸(G)铰链残基;3)磷酸丝氨酸(S)和/或磷酸苏氨酸(T)残基簇;以及4)高频率出现的带正电的精氨酸(R)和赖氨酸(K)以及带负电的天冬氨酸(D)和/或谷氨酸(E)氨基酸。另一方面,所提供的多肽包含羧基端的II型PDZ结合基序、与该PDZ结合基序相邻的脯氨酸(P)和/或甘氨酸(G)铰链残基以及与该铰链残基相邻的带正电残基(K、R、D、E)。
PDZ结构域最初被鉴定为突触后密度蛋白(postsynaptic densityprotein)PSD95/SAP90、果蝇(Drosophila)肿瘤抑制基因dlg-A和紧密连接蛋白(tight junction protein)ZO-1内的保守序列元件。尽管最初被称为GLGF或DHR基序,但是现在人们都把它们表示为PDZ,PDZ是最初发现含有该基序的三种蛋白质(PSD95/DLG/ZO-1)的首字母缩写。这些80-90氨基酸序列现已在75种以上蛋白质中得到鉴定,并在单一的蛋白质内就特征性地以多个拷贝得以表达。因此,一方面,所提供的多肽能抑制α连接蛋白结合包含PDZ结构域的蛋白质。PDZ结构域为一种特定类型的蛋白质相互作用构件(module),该构件具有结构非常明确、可由PDZ-结合基序(在本文中被称为“PDZ”基序)填充的相互作用“袋”。PDZ基序通常但并非总是位于胞内羧基最末端的共有序列。已分出四类PDZ基序:I型(S/T-x-Φ)、II型(Φ-x-Φ)、III型(Ψ-x-Φ)以及IV型(D-x-V),其中x为任何氨基酸,Φ为疏水残基(V、I、L、A、G、W、C、M、F),Ψ为碱性亲水残基(H、R、K)(Songyang,Z.,et al.1997.Science 275,73-77)。因此,一方面,所提供的多肽包含II型PDZ结合基序。
已注意到α Cx37的羧基最末端的18个氨基酸的序列代表了ACT肽主题中的一个例外变体。该Cx37ACT样序列为GQKPPSRPSSSASKKQ*YV(SEQ IDNO:43)。因此,Cx37羧基端的四个氨基酸仅有一部分与II型PDZ结合结构域相符。与典型的II型PDZ结合结构域不同,Cx37在本来预计为疏水氨基酸的第二位具有一个中性Q*。因此,Cx37包含可能被称为II型PDZ结合结构域样序列的序列。尽管如此,Cx37严格保持着ACT肽构成的所有其他方面,包括与PDZ结合结构域样序列相邻的成簇丝氨酸残基、高频率出现的R和K残基和富含P的序列。考虑到该ACT样构成的保守性的整体水平与上面列出的其它>70α连接蛋白相一致,则可以理解,Cx37 ACT样羧基端以所提供的能力起作用。
为了比较,表2给出了β连接蛋白Cx26。Cx26不具有羧基端II型PDZ结合基序;不到30%的羧基最末端氨基酸由S、T、R、D或E残基构成;没有证据表明含P和G铰链残基簇的基序与II型PDZ结合基序或PDZ样基序相邻;并且没有证据表明存在丝氨酸和苏氨酸磷酸化氨基酸的成簇重复样基序。Cx26的确有三个赖氨酸(K)残基,在所述序列羧基端附近彼此接连成簇。但是,在上文罗列的>70α连接蛋白中,并没有发现所研究的α连接蛋白呈现出在羧基端有三个重复K残基结构域这一特征(Cx26为β连接蛋白,因此在定义上就不含ACT结构域)。
本文提供的这段相对较短的氨基酸的独特功能性特征在减轻炎症、促进愈合、减少瘢痕形成、增加抗拉强度以及促进复杂组织结构再生中具有意料之外的作用,并且在不同组织如皮肤和脑损伤后起作用。因此,一方面,所提供多肽包含II型PDZ结合基序(Φ-x-Φ,其中x=任何氨基酸,Φ=疏水性氨基酸)。另一方面,所提供ACT多肽超过50%、60%、70%、80%、90%的氨基酸由一个或多个脯氨酸(P)、甘氨酸(G)、磷酸丝氨酸(S)、磷酸苏氨酸(T)、精氨酸(R)、赖氨酸(K)、天冬氨酸(D)或谷氨酸(E)氨基酸残基构成。
氨基酸脯氨酸(P)、甘氨酸(G)、精氨酸(R)、赖氨酸(K)、天冬氨酸(D)和谷氨酸(E)为蛋白质结构和功能的必需决定子。脯氨酸和甘氨酸残基可在蛋白质的3D结构中提供紧密转角,能形成功能所需的多肽的折叠构象。带电氨基酸序列通常位于折叠蛋白质的表面,是多肽介导的化学相互作用所必需的,所述化学相互作用包括蛋白质-蛋白质相互作用、蛋白质-脂质相互作用、酶-底物相互作用以及蛋白质-核酸相互作用。因此,另一方面,与II型PDZ结合基序相邻的脯氨酸(P)和甘氨酸(G)赖氨酸(K)、天冬氨酸(D)和谷氨酸(E)富集的区域可提供所提供的ACT肽的作用所必需的性质。另一方面,所提供的多肽包含与II型PDZ结合基序相邻的脯氨酸(P)和甘氨酸(G)赖氨酸(K)、天冬氨酸(D)和/或谷氨酸(E)富集的区域。
磷酸化是最常见的蛋白质翻译后修饰,并且在调整或修饰蛋白质结构和功能中至关重要。由磷酸化修饰的蛋白质结构和功能的方面包括蛋白质构象、蛋白质-蛋白质相互作用、蛋白质-脂质相互作用、蛋白质-核酸相互作用、通道门控、蛋白质运输和蛋白质转化。因此,一方面,磷酸丝氨酸(S)和/或磷酸苏氨酸(T)富集的序列对于改变ACT肽的功能、提高或降低多肽在其提供作用中的有效性是必需的。另一方面,所提供多肽包含丝氨酸(S)和/或磷酸苏氨酸(T)富集的序列或基序。
另一个实例中,鉴于ACT肽的定义,根据低等动物如鱼中较高程度的组织/器官再生潜能,甲硫氨酸幸而出现在斑马鱼Cx43的ACT序列的氨基端附近(表2)。除了编码甲硫氨酸,甲硫氨酸碱基对三联体为一个备选翻译起始位点。若翻译始于该甲硫氨酸,则可产生序列SSRARPDDLDV(SEQ IDNO:90)。该翻译产物保持标准ACT肽全部的保守和独特特征。具体而言,该肽包含羧基端II型PDZ结合结构域和与该PDZ结合结构域相邻的富含P、R和D残基的结构域。另外,该序列在其氨基端包含可能调节ACT肽功能的成簇S基序。这使人非常有兴趣地预期具有较高组织/器官再生潜能的动物如鱼可直接翻译ACT肽序列。
因此,所提供多肽包含人Cx43的羧基端序列。因此,所提供多肽可包含氨基酸序列SEQ ID NO:1或SEQ ID NO:2。该多肽可含有人Cx40羧基端的9个氨基酸。因此,该多肽包含氨基酸序列SEQ ID NO:5。
本文提及具体蛋白质时,考虑了变体、衍生物和片段。蛋白质变体和衍生物为本领域的普通技术人员所熟知,并且可氨基酸序列修饰物。例如,氨基酸序列修饰物一般分为三类中的一类或一类以上:置换型变体、插入型变体或缺失型变体。插入包括氨基端和/或羧基端融合以及一个或多个氨基酸残基的序列内插入。插入通常为比氨基端或羧基端融合改变程度小的插入,例如,数量级为一至四个残基的插入。缺失的特征在于将一个或多个氨基酸残基从蛋白质序列上除去。这些变体通常通过如下方法制备:对编码蛋白质的DNA中的核苷酸进行位点特异性诱变,由此产生编码变体的DNA,随后在重组细胞培养物中表达该DNA。在序列已知的DNA上的预定位点进行置换突变的技术已为人熟知,包括例如M13引物诱变和PCR诱变。氨基酸置换通常为单个氨基酸的置换,但可同时出现在多个不同位点;插入的数量级通常为约1至10个氨基酸残基。缺失或插入优选在一对相邻的氨基酸进行,即缺失两个残基或插入两个残基。置换、缺失、插入或其任一组合可联合起来获得最终构建体。突变不可将序列置于读框之外,并且优选不形成可能产生二级mRNA结构的互补区域,除非mRNA二级结构中需要这样的变化。置换变体为其中至少一个残基被除去并且在其位置插入了不同的残基的变体。这类置换通常依据以下表3进行,并将其称为保守置换。
表3:氨基酸置换
例如,用另一个生物学和/或化学相似的氨基酸置换一个氨基酸为本领域的技术人员所熟知的保守置换。例如,保守置换可为用一个疏水性残基置换另一个疏水性残基,或者用一个极性残基置换另一个极性残基。置换可包括表3所示组合。每个明确公开的序列的保守置换变化形式均包含在本文提供的多肽内。
一般而言,保守置换对所得多肽的生物学活性影响甚微乃至没有。在一个具体实例中,保守置换为肽的氨基酸置换,该置换基本不影响肽的生物学功能。肽可包含一个或多个氨基酸置换,例如2-10个保守置换,2-5个保守置换,4-9个保守置换,例如2、5或10个保守置换。
通过使用例如标准方法例如定向诱变或PCR对编码多肽的核苷酸序列进行操作,可产生得到含有一个或多个保守置换的多肽。或者,通过使用标准肽合成方法可产生得到含有一个或多个保守置换的多肽。可采用丙氨酸扫描(alanine scan)来鉴定蛋白质中的哪个氨基酸残基可允许氨基酸置换。在一个实例中,丙氨酸或其它保守氨基酸(例如下文罗列的那些氨基酸)置换一个或多个天然氨基酸时,该肽的生物学活性降低不超过25%,例如不超过20%,例如不超过10%。
关于保守置换的其他信息可参见Ben-Bassat et al.,(J.Bacteriol.169:751-7,1987),O′Regan et al.,(Gene 77:237-51,1989),Sahin-Tothet al,(Protein Sci.3:240-7,1994),Hochuli et al.,(Bio/Technology6:1321-5,1988)以及遗传学和分子生物学的标准教材等。
可采用置换或缺失诱变来插入N-糖基化(Asn-X-Thr/Ser)或O-糖基化(Ser或Thr)位点。半胱氨酸或其它不稳定残基的缺失也可以是所需要的。潜在蛋白酶解位点例如Arg的缺失或置换可通过使其中一个碱性残基缺失或者用谷氨酰胺酰或组氨酰残基置换一个残基完成。
某些翻译后衍生作用是由于重组宿主细胞对表达多肽的作用所致。谷氨酰氨酰和天冬酰胺酰残基通常在翻译后脱去酰胺基成为相应的谷氨酰和天冬氨酰残基。或者,在温和的酸性环境中使这些残基脱去酰胺基。其它翻译后修饰包括脯氨酸和赖氨酸的羟基化,丝氨酰或苏氨酰残基的羟基的磷酸化,赖氨酸、精氨酸和组氨酸侧链o-氨基的甲基化(T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,San Francisco pp 79-86[1983]),N端胺的乙酰化以及某些情况下羧基端羧基的酰胺化。
所理解的是,可掺入到所公开组合物中的氨基酸和肽类似物有很多种。例如,有多种D型氨基酸或具有与表3所示氨基酸功能不同的取代基的氨基酸。公开了天然存在的肽的对立立体异构体以及肽类似物的立体异构体。通过使tRNA分子运载所选择的氨基酸,并设计遗传构建体,很容易将这些氨基酸掺入多肽链,所述构建体利用例如琥珀密码子以位点特异性方式将氨基酸类似物插入肽链中(Thorson et al.,Methods in Molec.Biol.77:43-13(1991),Zoller,Current Opinion in Biotechnology,3:348-354(1992);Ibba,Biotechnology & Genetic Enginerring Reviews13:197-216(1995),Cahill et al,TIBS,14(10):400-403(1989);Benner,TIB Tech,12:158-163(1994);Ibba and Hennecke,Bio/technology,12:678-682(1994),至少就涉及氨基酸类似物的内容通过援引将其全部纳入本文)。
可产生与多肽相似,但并不通过天然肽键连接的分子。例如,连接氨基酸或氨基酸类似物的键包括CH2NH--、--CH2S--、--CH2--CH2--、-CH=CH-(顺式和反式)、--COCH2--、--CH(OH)CH2--和--CHH2SO-(这些内容和其它内容可参见Spatola,A.F.in Chemistry and Biochemistry of Amino Acids,Peptides,and Proteins,B.Weinstein,eds.,Marcel Dekker,New York,p.267(1983);Spatola,A.F.,Vega Data(March 1983),Vol.1,Issue3,Peptide Backbone Modifications(综述);Morley,Trends Pharm Sci(1980)pp.463-468;Hudson,D.et al,Int J Pept Prot Res 14:177-185(1979)(-CH2NH-,CH2CH2-);Spatola et al.Life Sci 38:1243-1249(1986)(-CH H2-S);Hann J.Chem.Soc Perkin Trans.1307-314(1982)(-CH--CH-,顺式和反式);Almquist et al.J.Med.Chem.23:1392-1398(1980)(-COCH2-);Jennings-White et al.Tetrahedron Lett 23:2533(1982)(-COCH2--);Szelke et al.European Appln,EP 45665 CA(1982):97:39405(1982)(-CH(OH)CH2-);Holladay et al.Tetrahedron.Lett24:4401-4404(1983)(-C(OH)CH2-);以及Hruby Life Sci 31:189-199(1982)(-CH2-S-);每篇文献均通过援引纳入本文。所理解的是,肽类似物在连接原子(bond atom)之间具有一个以上的原子,例如b-丙氨酸、g-氨基丁酸等。
氨基酸类似物和肽类似物通常具有增强或所需的性质,例如生产更为经济,在化学上更为稳定、药理性质(例如半衰期、吸收、效能、有效性等)得到增强、特异性(如生物活性的广谱性)改变、抗原性降低、穿过生物屏障(如消化道、血管、血脑屏障)的能力增强等等。
D型氨基酸可用来形成更为稳定的肽,原因在于D型氨基酸不会被肽酶等识别。用同类D型氨基酸系统置换共有序列的一个或多个氨基酸(如用D-赖氨酸置换L-赖氨酸)可用来形成更为稳定的肽。半胱氨酸残基可用来环化或者将两个或多个肽连接在一起。这有利于将肽限制在特定构象。(Rizo and Gierasch Ann.Rev.Biochem.61:387(1992),通过援引纳入本文)。
因此,所提供多肽可包含α连接蛋白c端(ACT)的保守变体。如表4所示,序列SEQ ID NO:2内的单个保守置换的一个实例在序列SEQ ID NO:3中给出。序列SEQ ID NO:2内的三个保守置换的一个实例在序列SEQ ID NO:4中给出。因此,所提供多肽包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4。
表4.ACT多肽变体
所理解的是,对本文公开的基因和蛋白质的任何变体、修饰物或衍生物进行定义的一种方法是通过根据与特定已知序列的同一性(在本文中也称为同源性)来定义变体、修饰物和衍生物。具体公开了本文公开的核酸和多肽的变体,该变体与所述序列或已知序列有至少百分之65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99的序列同一性。本领域的技术人员很容易知晓如何确定两种蛋白质或核酸的序列同一性。例如,可在对两个序列进行比对之后计算序列同一性,由此获得的序列同一性水平最高。
另一种计算序列同一性的方法可通过已发表的算法进行。对用于比较的序列的最佳比对可通过如下方法进行:Smith and Waterman Adv.Appl.Math.2:482(1981)的局部序列同一性算法,Needleman and Wunsch,J.MoL Biol.48:443(1970)的序列同一性比对算法,Pearson and Lipman,Proc.Natl.Acad.Sci.U.S.A.85:2444(1988)检索相似性的方法,这些算法的计算机执行(Wisconsin Genetics软件包的GAP、BESTFIT、FASTA和TFASTA,Genetics Computer Group,575 Science Dr.,Madison,WI)或检查。就计算序列同一性的方法将这些参考文献全文纳入本文。
核酸的同类序列同一性可由例如以下文献公开的算法获得:Zuker,M.Science 244:48-52,1989,Jaeger et al.Proc.Natl.Acad.Sci.USA86:7706-7710,1989,Jaeger et al.Methods Enzymol.183:281-306,1989至少就与核酸比对相关的内容将这些文献纳入本文。
因此,所提供多肽可包含与α连接蛋白c端(ACT)有至少百分之65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99序列同一性的氨基酸序列。因此,一方面,所提供多肽包含与SEQ IDNO:1、SEQ ID NO:29、SEQ ID NO:30、SEQ ID NO:31、SEQ IDNO:32、SFQ IDNO:33、SEQ ID NO:34、SEQ ID NO:35、SEQ ID NO:36、SEQ ID NO:37、SEQID NO:38、SEQ ID NO:39、SEQ ID NO:40、SEQ ID NO:41、SEQ ID NO:90或SEQ ID NO:91有至少百分之65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99序列同一性的氨基酸序列。作为一个实例,提供了与存在于人Cx43羧基端的同样是9个氨基酸的序列(SEQ ID NO:2)有66%序列同一性的多肽(SEQ ID NO:4)。
本文提供的多肽可直接添加至受试者的组织损伤处。但是,所提供多肽的胞质定位效率通过以化学方式与该肽顺式或反式连接的细胞内化运载体(cellular internalization transporter)得到增强。细胞内化运载体的效率还通过光照或与Tat-HA肽一起的细胞共转导得到增强。
因此,所提供多肽可包含细胞内化运载体或序列。细胞内化序列可为本领域已知或新发现的任何内化序列或其保守变体。细胞内化运载体和序列的非限制性实例包括触角足蛋白(Antennapedia)序列、TAT、HIV-Tat、穿膜肽(Penetrat in)、Antp-3A(Antp突变体)、Buforin II、Transportan、MAP(模式两亲肽(model amphipathic))、K-FGF、Ku70、朊病毒、pVEC、Pep-1、SynBl、Pep-7、HN-1、BGSC(双-胍-亚精胺-胆固醇)和BGTC(双-胍-Tren-胆固醇)(参见表5)。
表5:细胞内化运载体
因此,所提供的多肽可进一步包含氨基酸序列SEQ ID NO:7、SEQ IDNO:14(Bucci,M.et al 2000.Nat.Med 6,1362-1367)、SEQ ID NO:15(Derossi,D.,et al.1994.Biol.Chem.269,10444-10450)、SEQ ID NO:16(Fi scher,P.M.et al.2000.J.Pept.Res.55,163-172)、SEQ ID NO:17(Frankel,A.D.&Pabo,C.O.1988.Cell 55,1189-1193;Green,M.&Loewenstein,P.M.1988.Cell 55,1179-1188)、SEQ ID NO:18(Park,C.B.,et al.2000.Proc.Natl Acad.Sci.USA 97,8245-8250)、SEQID NO:19(Pooga,M.,et al.1998.FASEB J.12,61-11)、SEQ ID NO:20(Oehlke,J.et al.1998.Biochim.Biophys.Acta.1414,127-139)、SEQ ID NO:21(Lin,Y.Z.,et al.1995.J.Biol.Chem.270,14255-14258)、SEQ ID NO:22(Sawada,M.,et al.2003.Nature Cell Biol.5,352-357)、SEQ ID NO:23(Lundberg,P.et al.2002.Biochem.Biophys.Res.Commun.299,85-90)、SEQ ID NO:24(Elmquist,A.,et al.2001.Exp.Cell Res.269,237-244)、SEQ ID NO:25(Morris,M.C,et al.2001.Nature Biotechnol.19,1173-1176)、SEQ ID NO:26(Rousselle,C.et al.2000.Mol Pharmacol.57,679-686)、SEQ ID NO:27(Gao,C.et al.2002.Bioorg.Med.Chem.10,4057-4065)或SEQ ID NO:28(Hong,F.D.&Clayman,G.L.2000.Cancer Res.60,6551-6556)。所提供多肽可进一步包含BGSC(双-胍-亚精胺-胆固醇)或BGTC(双-胍-Tren-胆固醇)(Vigneron,J.P.et al.1998.Proc.Natl.Acad.Sci.USA.93,9682-9686)。就对细胞内化载体和序列的教导通过援引将上述参考文献全文纳入本文。现已知的或随后鉴定的任何其它内化序列可与本发明的肽结合。
所提供的多肽可包含与任何一个本文提供的细胞内化序列合的任何ACT序列(如任何一种本文公开的ACT肽)。所述结合的实例在表6中给出。因此,所提供的多肽可包含氨基酸序列SEQ ID NO:8、SEQ ID NO:9、SEQ IDNO:10、SEQ ID NO:11或SEQ ID NO:12。
表6:连有细胞内化序列(CIS)的ACT多肽
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。
![]()














动态评分
0.0