









专利摘要
本发明提供一株重组解脂耶罗维亚酵母及其构建方法和应用,涉及生物工程领域。该重组解脂耶罗维亚酵母,是解脂耶罗维亚酵母(Yarrowialipolytica)XJ‑8株,已在中国微生物菌种保藏管理委员会普通微生物中心保藏,保藏编号为CGMCCNo.19064。本发明重组解脂耶罗维亚酵母的构建方法,操作简单、高效,能够高效合成β‑榄香烯和/或大根香叶烯A。
权利要求
1.一种重组解脂耶罗维亚酵母,是解脂耶罗维亚酵母(
2.根据权利要求1重组解脂耶罗维亚酵母,其特征在于所述重组解脂耶罗维亚酵母(
3.根据权利要求2所述重组解脂耶罗维亚酵母,其特征在于所述大根香叶烯A合成酶来源于罗马洋甘菊(
4.根据权利要求3所述重组解脂耶罗维亚酵母,其特征在于所述重组解脂耶罗维亚酵母是在解脂耶罗维亚酵母基因组中插入了3-羟基-3-甲基戊二酰CoA还原酶表达盒和融合蛋白表达盒后所得;所述融合蛋白是由大根香叶烯A合成酶、连接肽和法尼基焦磷酸合成酶组成的;所述连接肽为GGGGS或GSG或EAAAK。
5.根据权利要求4所述重组解脂耶罗维亚酵母,其特征在于所述表达盒的启动子为解脂耶罗维亚酵母的TEFin启动子或TEF1启动子,终止子为解脂耶罗维亚酵母的xpr2t或cyc1终止子。
6.根据权利要求5所述重组解脂耶罗维亚酵母,其特征在于所述重组解脂耶罗维亚酵母还表达1个或多个标记基因;所述标记基因选自3(β)-异丙基苹果酸脱氢酶编码基因表达盒或乳清酸核苷-5′-磷酸脱羧酶编码基因表达盒。
7.根据权利要求6所述重组解脂耶罗维亚酵母,其特征在于所述大根香叶烯A合成酶的编码基因如SEQ ID No:1所示;所述3-羟基-3-甲基戊二酰CoA还原酶的编码基因序列如SEQID No:2所示。
8.权利要求1-7中任一所述重组菌在生产β-榄香烯和/或大根香叶烯A中的应用,其特征在于包括如下步骤:(1)采用发酵培养基培养权利要求1-7中任一所述重组菌,得到发酵产物;(2)有机溶液萃取所述发酵产物,收集有机相,得到大根香叶烯A;(3)将所述有机相加热到250-300℃,得到β-榄香烯。
说明书
技术领域
本发明涉及生物工程领域,尤其涉及生产β-榄香烯/或大根香叶烯A的重组解脂耶罗维亚酵母及其构建方法和应用。
背景技术
β-榄香烯(β-elemene),学名:1-甲基-1-乙烯基-2,4-二异丙基环己烷,是一种挥发性倍半萜,主要存在于莪术(Curcuma zedoaria)、香茅(Cymbopogon citratus)等植物的精油中。β-榄香烯能通过血脑屏障,不仅能直接抗肿瘤,而且还有免疫保护作用;同时具有缓解癌性疼痛,升高白细胞和抑制血小板聚集等特点(Edris AE,Current ClinicalPharmacology,2009,4(1):43-46.;Chen M et al.,Evidence-Based Complementary andAlternative Medicine,2012,2012.)。并且,与大多数化疗药物相比,该药毒副作用小,无明显肝、肾功能损害,不发生骨髓抑制,长期使用无耐药性(Zhai B et al.,InternationalJournal of Nanomedicine,2018,13:6279)。β-榄香烯是我国第一个得到国际认可,并具有自主知识产权的抗肿瘤药物。其口服乳和注射液作为国家二类新药在临床上已经取得了显著的疗效。
β-榄香烯主要是通过水蒸汽蒸馏和超临界CO2萃取等方法从莪术挥发油中分离提取获得;但由于莪术中β-榄香烯含量低,导致提取法投料多、耗时长、纯度和得率低而且步骤繁琐,不能完全适应市场的需求。并且植物提取的榄香烯具有多种异构体,提纯获得单一构型的β-榄香烯比较困难。由于微生物生长周期较短,可利用糖类原料在短时间内实现多批次生产,并且生物体内的酶催化可以获得单一构型的物质,相对于植物提取法而言,用微生物来制造β-榄香烯是一种经济高效的方式。
大根香叶烯A合成酶(GAS)能催化法尼基焦磷酸(FPP)合成大根香叶烯A,而大根香叶烯A在250℃下会发生Cope重排转化为β-榄香烯(GooréStéphane G et al.,NaturalProduct Research,2017,31(15):1836-1839.)。目前,已经有相关研究利用这一点在微生物中进行β-榄香烯的生产,但由于产量低未能实现工业化应用。
发明内容
本发明的目的是提供一种重组解脂耶罗维亚酵母,能够高效合成β-榄香烯和/或大根香叶烯A。
本发明再一目的是提供所述重组解脂耶罗维亚酵母的构建方法,该方法高效,操作简单。
本发明的再一目的是提供所述重组解脂耶罗维亚酵母在生产β-榄香烯和/或大根香叶烯A中的应用。
本发明的目的采用如下技术方案实现:
一种重组解脂耶罗维亚酵母,是解脂耶罗维亚酵母(Yarrowia lipolytica)XJ-8株,已在中国微生物菌种保藏管理委员会普通微生物中心保藏,保藏编号为CGMCCNo.19064。
在本发明中,所述重组解脂耶罗维亚酵母(Yarrowia lipolytica)XJ-8株是在解脂耶罗维亚酵母基因组中插入了大根香叶烯A合成酶、法尼基焦磷酸合成酶和3-羟基-3-甲基戊二酰CoA还原酶表达盒后得到的。
在本发明中,所述大根香叶烯A合成酶来源于罗马洋甘菊(Chamaemelum nobile),其编码基因经密码子优化后适合在解脂耶氏酵母解脂耶罗维亚酵母中表达,法尼基焦磷酸合成酶和3-羟基-3-甲基戊二酰CoA还原酶编码基因来源于解脂耶罗维亚酵母。
在本发明中,所述重组解脂耶罗维亚酵母是在解脂耶罗维亚酵母基因组中插入了3-羟基-3-甲基戊二酰CoA还原酶表达盒和融合蛋白表达盒后所得;所述融合蛋白是由大根香叶烯A合成酶、连接肽和法尼基焦磷酸合成酶(genbank登录号XP_503599.1)组成的;所述连接肽为GGGGS或GSG或EAAAK。
在本发明中,所述表达盒的启动子为解脂耶罗维亚酵母的TEFin启动子或TEF1启动子;所述终止子为解脂耶罗维亚酵母的xpr2t或cyc1终止子。
在本发明中,所述重组解脂耶罗维亚酵母还表达1个或多个标记基因;所述标记基因选自3(β)-异丙基苹果酸脱氢酶编码基因表达盒或乳清酸核苷-5′-磷酸脱羧酶编码基因表达盒。
在本发明中,所述大根香叶烯A合成酶的编码基因如SEQ ID No:1所示;所述3-羟基-3-甲基戊二酰CoA还原酶的编码基因序列如SEQ ID No:2所示。
在本发明中,包括将所述3-羟基-3-甲基戊二酰CoA还原酶表达盒、大根香叶烯A合成酶表达盒和法尼基焦磷酸合成酶表达盒通过质粒形式导入所述解脂耶氏酵母解脂耶罗维亚酵母,然后整合在解脂耶氏酵母解脂耶罗维亚酵母基因组上的步骤。
本发明还提供所述重组解脂耶罗维亚酵母的构建方法,包括将所述3-羟基-3-甲基戊二酰CoA还原酶表达盒和融合蛋白表达盒通过质粒形式导入所述解脂耶氏酵母解脂耶罗维亚酵母,然后整合在解脂耶氏酵母解脂耶罗维亚酵母基因组上的步骤;所述融合蛋白是由大根香叶烯A合成酶、连接肽和法尼基焦磷酸合成酶组成的。
本发明还提供所述重组菌在生产β-榄香烯和/或大根香叶烯A中的应用,其特征在于包括如下步骤:(1)采用发酵培养基培养权利要求1-7中任一所述重组菌,得到发酵产物;(2)有机溶液萃取所述发酵产物,收集有机相,得到大根香叶烯A;(3)将所述有机相加热到250-300℃,得到β-榄香烯。
本发明的重组解脂耶罗维亚酵母能够表达大根香叶烯A合成酶、过表达内源法尼基焦磷酸合成酶和3-羟基-3-甲基戊二酰CoA还原酶,实验证明该重组解脂耶罗维亚酵母能够高效发酵生产β-榄香烯和/或大根香叶烯A,实现了天然产物β-榄香烯和/或大根香叶烯A在解脂耶罗维亚酵母中的合成,本发明的重组解脂耶罗维亚酵母具有以下优点:
(1)本发明构建的重组解脂耶罗维亚酵母,是基于敲除了负责编码非同源重组基因ku70的解脂耶罗维亚酵母,使其同源重组能力增强,通过解脂耶罗维亚酵母自身的同源重组功能实现基因的整合,能大幅度提高导入基因的遗传稳定性。该重组解脂耶罗维亚酵母构建方法高效,操作简单。
(2)重组解脂耶罗维亚酵母通过过表达3-羟基-3-甲基戊二酰CoA还原酶编码基因tHMG、法尼基焦磷酸合成酶编码基因ERG20和大根香叶烯A合成酶编码基因gas,实现了β-榄香烯和/或大根香叶烯A的高效合成。
附图说明
图1是重组质粒pUC-leu-A08-Optgas的结构图,其中A08-up表示A08位点上游同源臂,A08-dm表示A08位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,leu表示3(β)-异丙基苹果酸脱氢酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图2显示了重组质粒pUC-HUH-IntC-tHMG的结构其中IntC-up表示IntC位点上游同源臂,IntC-dm表示IntC位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,URA表示乳清酸核苷-5’-磷酸脱羧酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图3显示了重组质粒pUC-HUH-SCP2-ERG20的结构,其中SCP2-up表示SCP2位点上游同源臂,SCP2-dm表示SCP2位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,URA表示乳清酸核苷-5’-磷酸脱羧酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图4是重组质粒pUC-leu-A08-Optgas-GSG-ERG20的结构图,其中A08-up表示A08位点上游同源臂,A08-dm表示A08位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,leu表示3(β)-异丙基苹果酸脱氢酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图5是重组质粒pUC-leu-A08-ERG20-GSG-Optgas的结构图,其中A08-up表示A08位点上游同源臂,A08-dm表示A08位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,leu表示3(β)-异丙基苹果酸脱氢酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图6是重组质粒pUC-leu-A08-ERG20-EAAAK-Optgas的结构图,其中A08-up表示A08位点上游同源臂,A08-dm表示A08位点下游同源臂,xpr2t表示终止子Txpr2t,TEFin表示启动子PTEFin,leu表示3(β)-异丙基苹果酸脱氢酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子和终止子)。
图7显示Yarrowia lipolytica XJ-8株(重组菌8)生产β-榄香烯的GC检测图。
具体实施方式
下面通过具体实施例对本发明作进一步的说明。
下面实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
解脂耶罗维亚酵母(Yarrowia lipolytica)Po1f,购于美国菌种保藏中心,编号为ATCC MYA-2613。
解脂耶罗维亚酵母(Yarrowia lipolytica)Po1fΔku70(MatA,△ku70::hisG,leu2-270,ura3-302,xpr2-322,axp1-2),缩写为Yarrowia lipolytica Po1fΔku70。Yarrowia lipolytica Po1fΔku70由解脂耶罗维亚酵母(Yarrowia lipolytica)Po1f敲除了负责非同源重组的编码基因ku70后构建得到(公开于Kretzschmar A et al.,Currentgenetics,2013,59(1-2):63-72.)。
A08位点整合质粒是将Yarrowia lipolytica Po1fΔku70基因组中染色体A上A08位点起始密码子上游2521bp、终止密码子下游2031bp的序列插入pUC57-leu载体(构建方法见实施例1)中所得,leu表达盒(包含Yarrowia lipolytica内源的启动子和终止子)在A08位点上下游序列之间。
IntC位点整合质粒是将Yarrowia lipolytica Po1fΔku70基因组中染色体C上IntC位点起始密码子上游1402bp和终止密码子下游1396bp的序列插入pUC57-hisG-ura-hisG(构建方法见实施例1)载体中所得,两个hisG标签编码基因在IntC位点上、下游序列之间。
SCP2位点整合质粒是将Yarrowia lipolytica Po1fΔku70基因组中染色体E上的SCP2位点起始密码子上游1523bp、终止密码子下游1524bp的序列插入pUC57-hisG-ura-hisG载体中所得,两个hisG标签编码基因在SCP2位点上、下游序列之间。
本发明中的引物序列见表2和表4-1、表4-2、表4-3和表4-4。
实施例1、基因元件的扩增与目标质粒的制备
(一)目标基因的制备
根据NCBI上提供的来自罗马洋甘菊的大根香叶烯A合成酶编码基因gas(genbank登录号KU589283.1)的核苷酸序列,经过密码子优化后,委托苏州金唯智生物科技有限公司合成优化后的大根香叶烯A合成酶编码基因Optgas,并插入质粒pUC57中,得到质粒pUC57-Optgas。Optgas的核苷酸序列如SEQ ID No:1所示。
根据NCBI上提供的Yarrowia lipolytica中3(β)-异丙基苹果酸脱氢酶编码基因leu的核苷酸序列(genbank登录号M37309.1),委托苏州金唯智生物科技有限公司合成基因leu,将3(β)-异丙基苹果酸脱氢酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子PTEFin、终止子Txpr2t)插入质粒pUC57中,得到质粒pUC57-leu。根据NCBI上提供的Yarrowialipolytica中的乳清酸核苷-5′-磷酸脱羧酶编码基因ura的核苷酸序列(genbank登录号AJ306421.1)和hisG标签(genbank登录号AF324729.1),委托苏州金唯智生物科技有限公司合成,将两个hisG标签编码基因序列插入质粒pUC57中,在两个hisG标签编码基因序列中插入乳清酸核苷-5′-磷酸脱羧酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子PTEFin、终止子Txpr2t),以便实现ura标记回收,得到质粒pUC57-hisG-ura-hisG。
以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,以tHMG-F和tHMG-R为引物扩增3-羟基-3-甲基戊二酰CoA还原酶编码基因tHMG。tHMG的核苷酸序列如SEQ ID No:2所示。
以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,以ERG20-F和ERG20-R为引物扩增法尼基焦磷酸合成酶编码基因ERG20(genbank登录号XP_503599.1)。
Yarrowia lipolytica内源的启动子PTEFin的核苷酸序列如SEQ ID No:3所示,Yarrowia lipolytica内源的终止子Txpr2t核苷酸序列如SEQ ID No:4所示。
(二)重组质粒的构建
重组质粒的结构见表1和图1-5。
1、重组质粒pUC-leu-A08-Optgas的构建
重组质粒pUC-leu-A08-Optgas是以pUC57-leu为骨架,插入了Yarrowialipolytica Po1fΔku70中A08位点起始密码子处上游同源臂A08-up和终止密码子下游同源臂A08-dm,在该上下游同源臂之间插入了Optgas基因表达盒(PTEFin-Optgas-Txpr2t),3(β)-异丙基苹果酸脱氢酶编码基因表达盒在该上下游同源臂之间,结构如图1。
以A08-TEFinp-F和TEFinp-Optgas-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增Optgas表达盒启动子PTEFin。以Optgas-xpr2t-F和A08-xpr2t-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板扩增Optgas表达盒终止子Txpr2t。
以质粒pUC57-Optgas为模板,以TEFinp-Optgas-F和Optgas-xpr2t-R为引物,扩增两端分别带有启动子PTEFin和终止子Txpr2t同源臂的Optgas基因。
上述PCR反应中所用PCR酶为宝日医生物技术(北京)有限公司的PrimeSTAR MaxDNA聚合酶。上述PCR扩增体系如下:
上述PCR的程序如下:98℃变性10s,55℃退火5s,72℃延伸(延伸时间=目标片段长度/1kb,单位min),重复30个循环。
用TaKaRa MiniBEST DNA Fragment Purification Kit(购自上海百赛生物技术股份有限公司)纯化回收各片段。
将A08位点整合质粒通过用NEB公司的限制性内切酶SnaB I酶切后,琼脂糖凝胶电泳胶回收线性化的A08位点整合质粒。
将线性化A08位点整合质粒和本实施例标题1构建的Optgas基因表达盒中的各元件(启动子PTEFin、Optgas片段和终止子Txpr2t)利用南京诺唯赞生物科技有限公司的ClonExpress MultiS One Step Cloning Kit实现一步克隆,反应体系见下表。将反应体系在37℃孵育30min后,得到环状的重组载体。
一步克隆的体系如下表:
其中,线性化载体(x)、插入片段(y)的使用量可由下面公式计算获得:每片段或线性化载体的最适使用量=[0.02×片段或线性化载体的碱基对数]ng。
将环状的重组载体转化大肠杆菌DH5α感受态细胞,通过氨苄抗性的平板筛选并通过菌落PCR及测序验证,得到阳性重组质粒pUC-leu-A08-Optgas。
2、重组质粒pUC-HUH-IntC-tHMG的构建
重组质粒pUC-HUH-IntC-tHMG是以pUC57-hisG-ura-hisG为骨架,插入了Yarrowialipolytica Po1fΔku70中IntC位点起始密码子上游同源臂和终止密码子下游同源臂,在该上下游同源臂之间插入了tHMG表达盒(启动子PTEFin、终止子Txpr2t),乳清酸核苷-5′-磷酸脱羧酶编码基因表达盒(包含Yarrowia lipolytica内源的启动子PTEFin、终止子Txpr2t)也在上下游同源臂之间,具体结构见图2。
以IntC-TEFinp-F和TEFinp-tHMG-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增tHMG表达盒启动子PTEFin。以tHMG-xpr2t-F和IntC-xpr2t-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增tHMG表达盒终止子Txpr2t。
以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,以TEFinp-tHMG-F和tHMG-xpr2t-R为引物扩增两端分别带有启动子PTEFin和终止子Txpr2t同源臂的tHMG基因。
用NEB公司的限制性内切酶Pac1对IntC位点整合质粒进行酶切后,琼脂糖凝胶电泳胶回收线性化IntC位点整合质粒。
将线性化IntC位点整合质粒和本实施例标题2构建的tHMG基因表达盒中的各元件(片段PTEFin、tHMG和Txpr2t)利用南京诺唯赞生物科技有限公司的ClonExpress MultiS OneStep Cloning Kit实现一步克隆,得到重组质粒pUC-HUH-IntC-tHMG。
3、重组质粒pUC-HUH-SCP2-ERG20的构建
重组质粒pUC-HUH-SCP2-ERG20是以pUC57-hisG-ura-hisG为骨架,插入了Yarrowia lipolytica Po1fΔku70中SCP2位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了ERG20表达盒(启动子PTEFin、终止子Txpr2t),乳清酸核苷-5′-磷酸脱羧酶编码基因表达盒也在上下游同源臂之间,具体结构见图3。
以SCP2-TEFinp-F和TEFinp-ERG20-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增ERG20表达盒启动子PTEFin。以ERG20-xpr2t-F和SCP2-xpr2t-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增ERG20表达盒终止子Txpr2t。
以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,以TEFinp-ERG20-F和ERG20-xpr2t-R为引物,扩增两端分别带有启动子PTEFin和终止子Txpr2t同源臂的ERG20基因。
用NEB公司的限制性内切酶Hind III对SCP2位点整合质粒进行酶切,琼脂糖凝胶电泳胶回收线性化SCP2位点整合质粒。
将线性化SCP2位点整合质粒和本实施例标题3构建的ERG20基因表达盒中的各元件(片段PTEFin、ERG20和Txpr2t)利用南京诺唯赞生物科技有限公司的ClonExpress MultiSOne Step Cloning Kit实现一步克隆,得到重组质粒pUC-HUH-SCP2-ERG20。
4、pUC-leu-A08-Optgas-GSG-ERG20的构建
重组质粒pUC-leu-A08-Optgas-GSG-ERG20是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70中A08位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白Optgas-GSG-ERG20的表达盒(PTEFin-Optgas-GSG-ERG20-Txpr2),3(β)-异丙基苹果酸脱氢酶编码基因表达盒也在该上下游同源臂之间。融合蛋白Optgas-GSG-ERG20的N端为大根香叶烯A合成酶,C端为法尼基焦磷酸合成酶,大根香叶烯A合成酶与法尼基焦磷酸合成酶之间以氨基酸序列为GSG的连接肽相连,具体结构见图4。
以A08-TEFinp-F和TEFinp-Optgas-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增Optgas-GSG-ERG20表达盒启动子PTEFin。以ERG20-xpr2t-F和A08-xpr2t-R为引物,以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,扩增Optgas-GSG-ERG20表达盒终止子Txpr2t。
以质粒pUC57-Optgas为模板,以TEFinp-Optgas-F和Optgas-ERG20-R为引物,扩增Optgas基因。
以Yarrowia lipolytica Po1fΔku70基因组DNA为模板,以Optgas-ERG20-F和ERG20-xpr2t-R为引物扩增ERG20基因。
A08位点整合质粒通过用NEB公司的限制性内切酶SnaB I酶切后,琼脂糖凝胶电泳胶回收线性化A08位点整合质粒。
将本实施例标题4扩增获得的Optgas-GSG-ERG20表达盒中各基因元件(片段PTEFin、Optgas基因、ERG20基因和Txpr2t)和线性化A08位点整合质粒利用南京诺唯赞生物科技有限公司的ClonExpress MultiS One Step Cloning Kit实现一步克隆,得到重组质粒pUC-leu-A08-Optgas-GSG-ERG20。
5、重组质粒pUC-leu-A08-ERG20-GSG-Optgas的构建
重组质粒pUC-leu-A08-ERG20-GSG-Optgas是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70的A08位点起始密码子上游同源臂和终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白ERG20-GSG-Optgas的表达盒(PTEFin-ERG20-GSG-Optgas-Txpr2t),3(β)-异丙基苹果酸脱氢酶编码基因表达盒也在该上下游同源臂之间。融合蛋白ERG20-GSG-Optgas的N端为法尼基焦磷酸合成酶,C端为大根香叶烯A合成酶,法尼基焦磷酸合成酶与大根香叶烯A合成酶之间以氨基酸序列为GSG的连接肽相连,具体结构见图5。
以A08-TEFinp-F和TEFinp-ERG20-R为引物,以Yarrowia lipolyticaΔku70基因组为模板,扩增ERG20-GSG-Optgas表达盒的启动子PTEFin。以Optgas-xpr2t-F和A08-xpr2t-R为引物,以Yarrowia lipolyticaΔku70基因组为模板,扩增ERG20-GSG-Optgas表达盒的终止子Txpr2t。
以Yarrowia lipolytica Po1fΔku70基因组为模板,以TEFinp-ERG20-F和ERG20-Optgas-R为引物,扩增ERG20基因。
以带有合成的Optgas基因的质粒pUC57-Optgas为模板,以ERG20-Optgas-F和Optgas-xpr2t-R为引物,扩增Optgas基因。
用NEB公司的限制性内切酶SnaB I酶切A08位点整合质粒,经琼脂糖凝胶电泳后胶回收线性化A08位点整合质粒。
将本实施例标题5扩增获得的ERG20-GSG-Optgas表达盒中各基因元件(片段PTEFin、ERG20基因、Optgas基因和Txpr2t)和线性化A08位点整合质粒,利用南京诺唯赞生物科技有限公司的ClonExpress MultiS One Step Cloning Kit实现一步克隆,得到重组质粒pUC-leu-A08-ERG20-GSG-Optgas。
表1各重组质粒中插入序列
表2引物序列
实施例2重组菌的构建
(一)重组菌1的构建
(1)将含有Optgas基因表达盒的质粒pUC-leu-A08-Optgas导入Yarrowialipolytica Po1fΔku70中,Optgas表达盒整合到基因组A08位点处,得到重组菌1。具体方法如下:(1)Yarrowia lipolytica Po1fΔku70于YPD液体培养基(含有2%蛋白胨、1%酵母提取物和2%葡萄糖)中过夜培养后制备感受态细胞。(2)利用Zymo Research Corporation的Zymogen Frozen EZ Yeast Transformation Kit II将pUC-leu-A08-Optgas转化Yarrowia lipolytica Po1fΔku70感受态细胞,进行同源重组。
(2)采用筛选培养基SD-Leu筛选,将PCR鉴定正确的阳性克隆,命名为重组菌1。其中筛选培养基SD-Leu含有:葡萄糖20g/L,Yeast Nitrogen Base(YNB,酵母氮碱硫酸铵,购自BBI Life Sciences)6.7g/L,CSM-Leu(完全补充混合物除去亮氨酸,购自MPBiomedicals)0.67g/L,琼脂粉23g/L。
(二)重组菌2的构建
将重组质粒pUC-HUH-IntC-tHMG和pUC-HUH-SCP2-ERG20导入重组菌1,tHMG表达盒整合至基因组IntC位点,ERG20表达盒整合至基因组SCP2位点,得到重组菌2。具体方法如下:将重组菌1于YPD液体培养基(含有2%蛋白胨,1%酵母提取物,2%葡萄糖)中过夜培养后制备感受态,利用Zymo Research Corporation的Zymogen Frozen EZ YeastTransformation Kit II将重组质粒pUC-HUH-IntC-tHMG和pUC-HUH-SCP2-ERG20转化入重组菌1中,进行同源重组,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆,命名为重组菌2。
其中筛选培养基SD-Leu-Ura的成分为:葡萄糖20g/L,YNB 6.7g/L,CSM-Leu-Ura(完全补充混合物除去亮氨酸和尿嘧啶,购自MP Biomedicals)0.67g/L,琼脂粉23g/L。
(三)重组菌3的构建
将pUC-HUH-IntC-tHMG和pUC-leu-A08-Optgas-GSG-ERG20质粒导入Yarrowialipolytica Po1fΔku70中,tHMG表达盒整合至基因组中IntC位点,Optgas-GSG-ERG20表达盒整合至A08位点,得到重组菌3。具体方法如下:将Yarrowia lipolytica Po1fΔku70于YPD液体培养基(含有2%蛋白胨,1%酵母提取物,2%葡萄糖)中过夜培养后制备感受态,利用Zymo Research Corporation的Zymogen Frozen EZ Yeast Transformation Kit II将pUC-HUH-IntC-tHMG和pUC-leu-A08-Optgas-GSG-ERG20转化Yarrowia lipolytica Po1fΔku70,进行同源重组,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆,命名为重组菌3。
筛选培养基SD-Leu-Ura的成分为:葡萄糖20g/L,YNB(酵母氮碱硫酸铵,购自BBILife Sciences)6.7g/L,CSM-Leu-Ura(完全补充混合物除去亮氨酸和尿嘧啶,购自MPBiomedicals)0.67g/L,琼脂粉23g/L。
(四)重组菌4的构建
将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-ERG20-GSG-Optgas导入Yarrowialipolytica Po1fΔku70中,tHMG表达盒整合至基因组中IntC位点,ERG20-GSG-Optgas表达盒整合至A08位点,得到重组菌4。
具体方法如下:将Yarrowia lipolytica Po1fΔku70于YPD液体培养基(2%蛋白胨,1%酵母提取物,2%葡萄糖)中过夜培养后制备感受态,利用Zymo ResearchCorporation的Zymogen Frozen EZ Yeast Transformation Kit II将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-ERG20-GSG-Optgas转化Yarrowia lipolytica Po1fΔku70,采用筛选培养基SD-Leu-Ura筛选,PCR鉴定出正确的阳性克隆,得到重组菌4。
其中筛选培养基SD-Leu-Ura的成分为:葡萄糖20g/L,YNB 6.7g/L,CSM-Leu-Ura0.67g/L,琼脂粉23g/L。
(五)重组菌5-8的构建
以质粒pUC57-Optgas和Yarrowia lipolytica Po1fΔku70基因组DNA为模板,用表4(包括表4-1、4-2、4-3、4-4)的引物分别进行PCR扩增,得到不同引物对应的含有不同同源臂的Optgas和ERG20片段;再分别将不同的引物对应的扩增产物进行一步克隆组成融合蛋白表达盒质粒,将该载体中融合蛋白所含两酶之间的连接肽GSG分别替换成GGGGS、EAAAK。将上述质粒同pUC-HUH-IntC-tHMG质粒转入Yarrowia lipolytica Po1fΔku70中进行同源重组,分别得到重组菌5-8。
以带有合成的Optgas基因的质粒pUC57-Optgas为模板,以表4(表4-1、4-2、4-3、4-4)中连接肽为GGGGS/EAAAK的各对引物Optgas-F和Optgas-R进行PCR扩增Optgas基因,得到对应于引物对Optgas-F(Optgas-GGGGS-ERG20)和Optgas-R(Optgas-GGGGS-ERG20)、引物对Optgas-F(Optgas-EAAAK-ERG20)和Optgas-R(Optgas-EAAAK-ERG20)、引物对Optgas-F(ERG20-GGGGS-Optgas)和Optgas-R(ERG20-GGGGS-Optgas)、引物对Optgas-F(ERG20-EAAAK-Optgas)和Optgas-R(ERG20-EAAAK-Optgas)的Optgas扩增片段。
以Yarrowia lipolytica Po1fΔku70基因组为模板,以表4(表4-1、4-2、4-3、4-4)中连接肽为GGGGS/EAAAK的各对应引物ERG20-F和ERG20-R进行PCR扩增ERG20基因,得到对应引物对ERG20-F(Optgas-GGGGS-ERG20)和ERG20-R(Optgas-GGGGS-ERG20)、引物对ERG20-F(Optgas-EAAAK-ERG20)和ERG20-R(Optgas-EAAAK-ERG20)、引物对ERG20-F(ERG20-GGGGS-Optgas)和ERG20-R(ERG20-GGGGS-Optgas)、引物对ERG20-F(ERG20-EAAAK-Optgas)和ERG20-R(ERG20-EAAAK-Optgas)扩增的ERG20片段。
利用一步克隆分别将不同引物对对应的Optgas扩增片段和ERG20扩增片段作为融合蛋白表达盒元件构建重组质粒pUC-leu-A08-Optgas-GGGGS-ERG20、pUC-leu-A08-Optgas-EAAAK-ERG20、pUC-leu-A08-ERG20-GGGGS-Optgas、pUC-leu-A08-ERG20-EAAAK-Optgas。其中各融合蛋白表达盒元件的启动子均为Yarrowia lipolytica内源启动子PTEFin,终止子均为Yarrowia lipolytica内源终止子Txpr2t。其中融合蛋白ERG20-EAAAK-Optgas的表达盒启动子PTEFin扩增引物为A08-TEFinp-F和TEFinp-Optgas-R,终止子Txpr2t扩增引物为ERG20-xpr2t-F和A08-xpr2t-R。以表4-4中引物扩增获得的ERG20、Optgas片段和PTEFin、Txpr2t为融合蛋白ERG20-EAAAK-Optgas原件。
重组质粒pUC-leu-A08-Optgas-GGGGS-ERG20是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70 A08位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白Optgas-GGGGS-ERG20的表达盒(PTEFin-Optgas-GGGGS-ERG20-Txpr2t),负责合成亮氨酸的3(β)-异丙基苹果酸脱氢酶编码基因表达盒也在该上下游同源臂之间。融合蛋白Optgas-GGGGS-ERG20的N端为大根香叶烯A合成酶,C端为法尼基焦磷酸合成酶,大根香叶烯A合成酶法与法尼基焦磷酸合成酶之间以氨基酸序列为GGGGS的连接肽相连。
重组质粒pUC-leu-A08-Optgas-EAAAK-ERG20是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70 A08位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白Optgas-EAAAK-ERG20的表达盒(PTEFin-Optgas-EAAAK-ERG20-Txpr2t),负责合成亮氨酸的3(β)-异丙基苹果酸脱氢酶编码基因表达盒在该上下游同源臂之间。融合蛋白Optgas-EAAAK-ERG20的N端为大根香叶烯A合成酶,C端为法尼基焦磷酸合成酶,大根香叶烯A合成酶与法尼基焦磷酸合成酶之间以氨基酸序列为EAAAK的连接肽相连。
重组质粒pUC-leu-A08-ERG20-GGGGS-Optgas是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70 A08位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白ERG20-GGGGS-Optgas的表达盒(PTEFin-ERG20-GGGGS-Optgas-Txpr2t),负责合成亮氨酸的3(β)-异丙基苹果酸脱氢酶编码基因表达盒在该上下游同源臂之间。融合蛋白ERG20-GGGGS-Optgas的N端为法尼基焦磷酸合成酶,C端为大根香叶烯A合成酶,法尼基焦磷酸合成酶与大根香叶烯A合成酶之间以氨基酸序列为GGGGS的连接肽相连。
重组质粒pUC-leu-A08-ERG20-EAAAK-Optgas是以pUC57-leu为骨架,插入了Yarrowia lipolytica Po1fΔku70 A08位点起始密码子上游同源臂、终止密码子下游同源臂,在该上下游同源臂之间插入了融合蛋白ERG20-EAAAK-Optgas的表达盒(PTEFin-ERG20-EAAAK-Optgas-Txpr2t,),负责合成亮氨酸的3(β)-异丙基苹果酸脱氢酶编码基因表达盒在该上下游同源臂之间。融合蛋白ERG20-EAAAK-Optgas的N端为法尼基焦磷酸合成酶,C端为大根香叶烯A合成酶,法尼基焦磷酸合成酶与大根香叶烯A合成酶之间以氨基酸序列为EAAAK的连接肽相连,具体结构见图6。
将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-Optgas-GGGGS-ERG20导入Yarrowia lipolytica Po1fΔku70中,进行同源重组,tHMG表达盒整合至基因组IntC位点处,Optgas-GGGGS-ERG20表达盒整合至基因组A08位点处,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆命名为重组菌5。
将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-Optgas-EAAAK-ERG20导入Yarrowia lipolytica Po1fΔku70中,进行同源重组,tHMG表达盒整合至基因组IntC位点处,Optgas-EAAAK-ERG20表达盒整合至基因组A08位点处,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆,命名为重组菌6。
将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-ERG20-GGGGS-Optgas导入Yarrowia lipolytica Po1fΔku70中,进行同源重组,tHMG表达盒整合至基因组IntC位点处,ERG20-GGGGS-Optgas表达盒整合至基因组A08位点处,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆,命名为重组菌7。
将重组质粒pUC-HUH-IntC-tHMG和pUC-leu-A08-ERG20-EAAAK-Optgas导入Yarrowia lipolytica Po1fΔku70中,进行同源重组,tHMG表达盒整合至基因组中IntC位点,ERG20-EAAAK-Optgas表达盒整合至A08位点,采用筛选培养基SD-Leu-Ura筛选,将PCR鉴定正确的阳性克隆,命名为重组菌8。
其中,重组菌8已于2019年12月02日保藏在中国微生物菌种保藏管理委员会普通微生物中心(CGMCC),地址:北京市朝阳区北辰西路1号院3号,分类命名:解脂耶罗维亚酵母Yarrowia lipolytica,参椐的生物材料(株):XJ-8,保藏编号:CGMCC No.19064。
表3为连接肽核苷酸序列和氨基酸序列
表4-1为构建重组质粒pUC-leu-A08-Optgas-GGGGS-ERG20所需引物
表4-2为构建重组质粒pUC-leu-A08-Optgas-EAAAK-ERG20所需引物
表4-3为构建重组质粒pUC-leu-A08-ERG20-GGGGS-Optgas所需引物
表4-4为构建重组质粒pUC-leu-A08-ERG20-EAAAK-Optgas所需引物
实施例3、重组菌1-8在生产β-榄香烯中的应用
1、工程菌的培养及产物提取
分别采用实施例2中重组菌1-8生产β-榄香烯。具体方法如下:活化重组菌,于YPD液体培养基中30℃、220rpm条件下培养16h,得到种子液。将种子液以1%的接种量接种于50ml发酵培养基中,在30℃、220rpm震荡培养1天,然后加入发酵液体积10%的正十二烷,继续震荡培养3天。发酵结束后,将发酵液转移至50ml离心管,5000rpm离心15min,收集有机相备用。
其中发酵培养基含有55g/L葡萄糖、10g/L酵母提取物和20g/L胰蛋白胨。
2、β-榄香烯的定性定量分析
1)β-榄香烯转化
将上述有机相样品在通风厨内经220℃油浴加热1小时除去正十二烷,得到各重组菌转化后的物质。
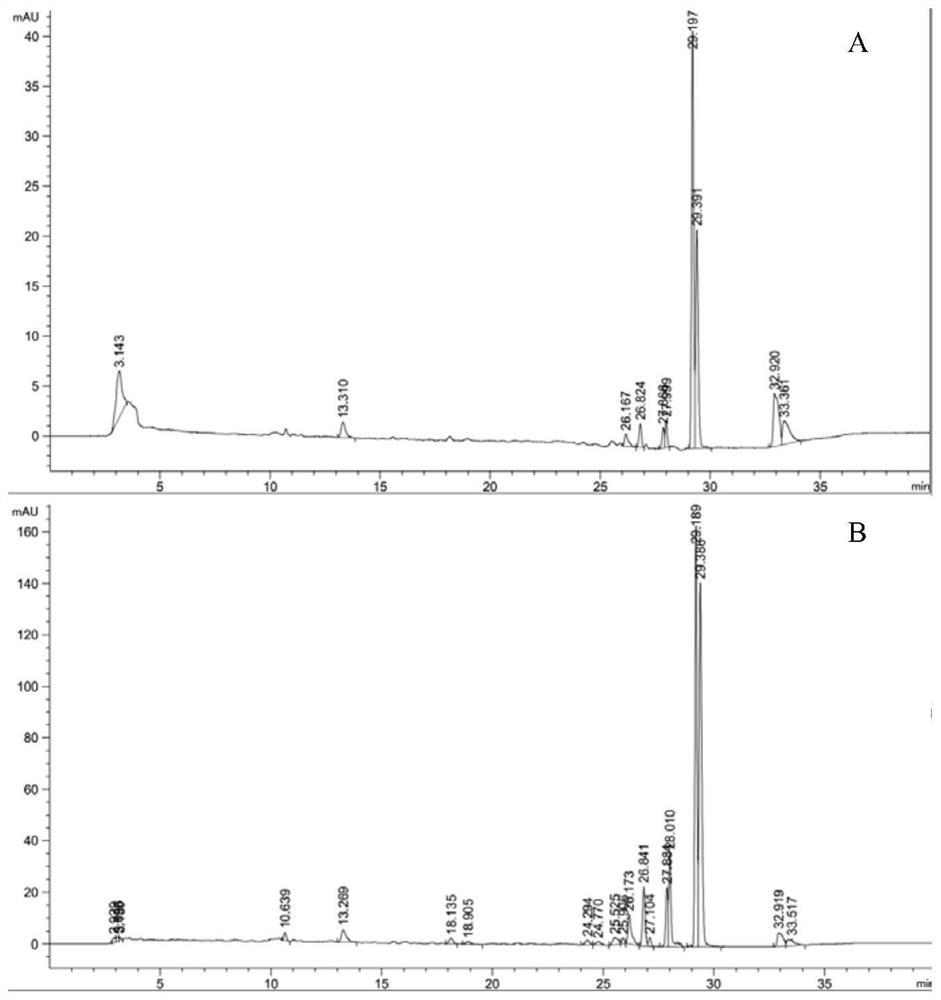
2)β-榄香烯检测
将各重组菌转化后的物质用正己烷稀释10倍后过有机尼龙滤膜(0.22um),用GC检测。检测条件:进样口温度250℃,进样体积1ul,不分流;色谱柱:HP-5ms(30m*0.25mM);色谱条件:初始温度为60℃,按照10℃/min的速度上升到160℃,然后5℃/min上升到200℃,最后10℃/min上升到230℃。用β-榄香烯的标准品进行定性定量。
图7为重组菌8发酵2天后生产的β-榄香烯的GC检测图。发酵5天后,重组菌8的β-榄香烯产量最高,达到了1052mg/L,即每升发酵液产1052mg的β-榄香烯。
重组菌1、重组菌2、重组菌3、重组菌4、重组菌5、重组菌6、重组菌7产量分别为61mg/L、201mg/L、254mg/L、187mg/L、79mg/L、246mg/L和413mg/L。
SEQUENCE LISTING
<110> 南京工业大学
<120> 一株重组解脂耶罗维亚酵母及其构建方法和应用
<130> 20200121
<160> 4
<170> PatentIn version 3.3
<210> 1
<211> 1683
<212> DNA
<213> 罗马洋甘菊
<400> 1
atggccgctg tccaagccaa cgtcaccacc ggcgtccaag ccaacaccaa gacctctgcc 60
aagcccgtca gacctctggc caacttcccc ccttccgtgt ggggtaccag attcctctcc 120
ttctctctgg acaagtccga gttcgagcga tatgccatgg ccatggaaaa gcccaaggag 180
gatctgcgaa atctgattgt ggaccccaag atggactcca acgaaaagct cggtctgatc 240
tactccgtgc atcgactcgg tctgacctac attttcctcg aggagatcga gtctcagctg 300
gacaagctct tcaacgagtt ctctctgcaa gactacgaag acgtggatct gtacaccatc 360
tctatcaact tccaagtctt tcgacatctg ggctacaagc tcgcttgcga cgtgttcaat 420
aaattcaagg acgtgtcctc cggcaccttc aaagagtcca ttacctccga cgtccgaggt 480
atgctgggtc tgtacgagtc tgcccagctg cggatccgag gcgaaaagat tctggacgag 540
gcttccgtgt tcatcgaggg taagctgaag tctgtcgtga acactctgga gtgcaatctc 600
gcccagcaag tcaagcagtc tctgaagcga cccttccacc aaggcatgcc cattgtggag 660
gcccgactct acttttccaa ctacgaggag gaatgttcct cccacgactc tctgttcaag 720
ctcgccaaac tgcacttcaa gtatctggag ctccagcaga aggaggagct ccgaatcgtg 780
accaagtggt acaaggacat gcgattccaa gaggacaccc cttacgtccg agatcgagtc 840
cccgagatct atctctggat cctcggcctc tactttgagc cccattactc tctcgctcga 900
atcatcgcca ccaagatcac tctgtttctc ctcatcctcg atgacaccta cgacgcttac 960
gccaccatcg acgagattcg actgctgact tatgccatta ataagtggga catttccgct 1020
atggagcaga tccccgaata catccgaccc ttttacaaag tcctcatcaa cgagtacgcc 1080
gagatcgaga agaagatggc taaggagggt cgagccaaca ccgtgatcgc ctccaaggag 1140
gccttccaag acattgctcg aggccatctg gaagaggccg agtggaccaa ctctggttat 1200
gtggcctcct tccccgacta catgaagaac ggtctgatta cctccggcta caacctcatc 1260
tccaagtctg ctctggtggg catgggtgac aacgtcggtg aggacgctct ggcttggtac 1320
gagtctcacc ccaagcctct gcaagcctcc gaactgattt ccagactgca agacgacgtg 1380
atgacctacc agttcgaacg agaacgaggt cagtctgcca cctccgtcga cgcctatatc 1440
aagacctacg gcgtgtccga aaagaaagcc attgacgagc tgaagtccat gatcgagaat 1500
gcttggaagg acatcaacga gggctgtctc aagccccgac aagtccctat ggatctgctg 1560
gccatcattc tgaacctcgc ccgaatgatc gacgtgatct accgatacga cgacggtttc 1620
accttccaag gctctaccat gaaggagtac attaacattc tgttcgtcga ctctctgccc 1680
gtg 1683
<210> 2
<211> 1503
<212> DNA
<213> 解脂耶罗维亚酵母
<400> 2
atgacccagt ctgtgaaggt ggttgagaag cacgttccta tcgtcattga gaagcccagc 60
gagaaggagg aggacacctc ttctgaagac tccattgagc tgactgtcgg aaagcagccc 120
aagcccgtga ccgagacccg ttctctggac gacctagagg ctatcatgaa ggcaggtaag 180
accaagcttc tggaggacca cgaggttgtc aagctctctc tcgagggcaa gcttcctttg 240
tatgctcttg agaagcagct tggtgacaac acccgagctg ttggcatccg acgatctatc 300
atctcccagc agtctaatac caagacttta gagacctcaa agcttcctta cctgcactac 360
gactacgacc gtgtttttgg agcctgttgc gagaacgtta ttggttacat gcctctcccc 420
gttggtgttg ctggccccat gaacattgat ggcaagaact accacattcc tatggccacc 480
actgagggtt gtcttgttgc ctcaaccatg cgaggttgca aggccatcaa cgccggtggc 540
ggtgttacca ctgtgcttac tcaggacggt atgacacgag gtccttgtgt ttccttcccc 600
tctctcaagc gggctggagc cgctaagatc tggcttgatt ccgaggaggg tctcaagtcc 660
atgcgaaagg ccttcaactc cacctctcga tttgctcgtc tccagtctct tcactctacc 720
cttgctggta acctgctgtt tattcgattc cgaaccacca ctggtgatgc catgggcatg 780
aacatgatct ccaagggcgt cgaacactct ctggccgtca tggtcaagga gtacggcttc 840
cctgatatgg acattgtgtc tgtctcgggt aactactgca ctgacaagaa gcccgcagcg 900
atcaactgga tcgaaggccg aggcaagagt gttgttgccg aagccaccat ccctgctcac 960
attgtcaagt ctgttctcaa aagtgaggtt gacgctcttg ttgagctcaa catcagcaag 1020
aatctgatcg gtagtgccat ggctggctct gtgggaggtt tcaatgcaca cgccgcaaac 1080
ctggtgaccg ccatctacct tgccactggc caggatcctg ctcagaatgt cgagtcttcc 1140
aactgcatca cgctgatgag caacgtcgac ggtaacctgc tcatctccgt ttccatgcct 1200
tctatcgagg tcggtaccat tggtggaggt actattttgg agccccaggg ggctatgctg 1260
gagatgcttg gcgtgcgagg tcctcacatc gagacccccg gtgccaacgc ccaacagctt 1320
gctcgcatca ttgcttctgg agttcttgca gcggagcttt cgctgtgttc tgctcttgct 1380
gccggccatc ttgtgcaaag tcatatgacc cacaaccggt cccaggctcc tactccggcc 1440
aagcagtctc aggccgatct gcagcgtcta caaaacggtt cgaatatttg catacggtca 1500
tag 1503
<210> 3
<211> 531
<212> DNA
<213> 解脂耶罗维亚酵母
<400> 3
agagaccggg ttggcggcgc atttgtgtcc caaaaaacag ccccaattgc cccaattgac 60
cccaaattga cccagtagcg ggcccaaccc cggcgagagc ccccttcacc ccacatatca 120
aacctccccc ggttcccaca cttgccgtta agggcgtagg gtactgcagt ctggaatcta 180
cgcttgttca gactttgtac tagtttcttt gtctggccat ccgggtaacc catgccggac 240
gcaaaataga ctactgaaaa tttttttgct ttgtggttgg gactttagcc aagggtataa 300
aagaccaccg tccccgaatt acctttcctc ttcttttctc tctctccttg tcaactcaca 360
cccgaaatcg ttaagcattt ccttctgagt ataagaatca ttcaaaatgg tgagtttcag 420
aggcagcagc aattgccacg ggctttgagc acacggccgg gtgtggtccc attcccatcg 480
acacaagacg ccacgtcatc cgaccagcac tttttgcagt actaaccgca g 531
<210> 4
<211> 516
<212> DNA
<213> 解脂耶罗维亚酵母
<400> 4
gatccaacta cggaacttgt gttgatgtct ttgcccccgg ctccgatatc atctctgcct 60
cttaccagtc cgactctggt actttggtct actccggtac ctccatggcc tgtccccacg 120
ttgccggtct tgcctcctac tacctgtcca tcaatgacga ggttctcacc cctgcccagg 180
tcgaggctct tattactgag tccaacaccg gtgttcttcc caccaccaac ctcaagggct 240
ctcccaacgc tgttgcctac aacggtgttg g
一株重组解脂耶罗维亚酵母及其构建方法和应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0