

IPC分类号 : C12N15/53I,C12N15/81I,C12N1/19I,C12N9/04I,C12Q1/00I,C12R1/84N






专利摘要
本发明提供了一种密码子优化的葡萄糖氧化酶基因,所述基因来源于AspergillusnigerAn76,其序列如SEQIDNo.2所示。该原始的葡萄糖氧化酶基因在毕赤酵母中无法成功表达,经过序列优化,其能够在毕赤酵母中顺利表达,并且,经过分离、纯化,能够得到纯化的目的蛋白,该重组葡萄糖氧化酶可以用于葡萄糖生物传感器中。
权利要求
1. 一种密码子优化的葡萄糖氧化酶基因,其特征在于,所述基因的核苷酸序列如SEQID No.2所示。
2.包含权利要求1所述基因的重组载体,所述载体骨架为pPIC9K载体。
3.包含权利要求2所述重组载体的重组菌株。
4.根据权利要求3所述的重组菌株,其特征在于,所述菌株为大肠杆菌或毕赤酵母。
5.根据权利要求4所述的重组菌株,其特征在于,所述大肠杆菌为大肠杆菌DH5α。
6.根据权利要求4所述的重组菌株,其特征在于,所述毕赤酵母为毕赤酵母GS115。
7.一种制备重组葡萄糖氧化酶的方法,其特征在于,所述方法包括如下步骤:
将权利要求1所述的基因转化到毕赤酵母中,构建重组表达葡萄糖氧化酶的毕赤酵母,然后,对重组的毕赤酵母进行培养,并对重组的葡萄糖氧化酶进行纯化。
8.根据权利要求7所述的方法,其特征在于,所述毕赤酵母为毕赤酵母GS115。
9.权利要求1所述的基因在制备葡萄糖氧化酶电极中的应用。
10.根据权利要求9所述的应用,其特征在于,所述葡萄糖氧化酶电极为将葡萄糖氧化酶固定在修饰的金电极上得到的葡萄糖氧化酶电极。
11.根据权利要求10所述的应用,其特征在于,所述修饰的金电极为采用半胱胺和纳米金修饰的金电极。
12.权利要求1所述的基因在制备葡萄糖生物传感器中的应用。
说明书
技术领域
本发明涉及生物酶基因工程和生物传感技术领域,具体涉及一种密码子优化的葡萄糖氧化酶基因及其在制备生物传感元件中的应用。
背景技术
目前,我国生物发酵产品总量居世界第一位,其中葡萄糖是发酵产品重要的质量控制指标,快速、准确检测发酵产品中的葡萄糖含量对于保证发酵产品质量具有重要意义。发酵产品中葡萄糖的传统检测方法主要包括高效液相色谱(HPLC)、气相色谱(GC)、旋光以及薄层色谱法,但是这些检测方法面临分析成本高、操作繁琐等各种问题。而生物传感器法具有灵敏度高、重现性好、操作简单的特点而受到广泛青睐。
葡萄糖氧化酶(Glucose oxidase(GOD))是葡萄糖生物传感器中主要的检测敏感元件,GOD是一种黄素蛋白,以氧气为电子受体催化β-D-葡萄糖氧化为D-葡萄糖酸-δ-内酯和H2O2,目前我国GOD主要分离纯化自黑曲霉或青霉,这种来源的酶产量很低而且纯化过程繁琐,这就导致生产的葡萄糖氧化酶远远不能满足应用需求,大部分商品化酶的来源依赖Sigma等国外公司,作为酶电极的“微芯片”,长期依赖进口,容易导致“卡脖子”的问题。
葡萄糖氧化酶活性中心深埋在酶分子内部,酶分子与电极之间很难进行直接电子传递,目前研究较多的是利用具有导电性高、表面活性中心多、生物相容性好等特殊性质的纳米材料作为电子导线介导酶与电极之间的电子传递,许多纳米材料,如纳米金属、纳米氧化物以及碳纳米管,已经用于葡萄糖氧化酶传感器中酶电极传感器的制作,并使传感器的传感性能得到明显提升。但是这些传感器的酶负载量、灵敏度、检测范围、响应时间等有待进一步改进,因此需要进一步研究葡萄糖氧化酶、纳米材料与电极之间的直接电化学。
毕赤酵母具有遗传背景清晰、外源蛋白表达量高、可对蛋白进行正确折叠、修饰和加工、可进行高密度发酵等优势,因此可作为外源蛋白的优良表达宿主。但是,将不同来源的葡萄糖氧化酶在毕赤酵母中进行表达时,通常会遇到无法成功表达的情况,需要对葡萄糖氧化酶的基因序列进行优化;对基因序列进行优化的方式多种多样,如何调整基因序列并使其在毕赤酵母中顺利表达是很难预期的。
发明内容
本发明提供了一种密码子优化的葡萄糖氧化酶。
本发明提供的密码子优化的葡萄糖氧化酶基因来源于AspergillusnigerAn76,其原始的基因序列如SEQ ID No.1所示,经过序列优化的基因序列如SEQ ID No.2所示。该原始的葡萄糖氧化酶基因在毕赤酵母中无法成功表达,经过序列优化,其能够在毕赤酵母中顺利表达,并且,经过分离、纯化,能够得到纯化的目的蛋白。
另一方面,本发明还提供了包含葡萄糖氧化酶基因的重组载体。本发明所述的载体包括克隆载体以及表达载体。
在一个实施方案中,所述克隆载体包括pUC系列的载体,例如,pUC18、pUC19;所述表达载体包括在大肠杆菌中表达的载体以及在毕赤酵母中表达的载体,优选的,包括pET系列的载体,如pET-21a;更优选的,所述在毕赤酵母中表达的载体为pPIC9K载体。
另一方面,本发明还提供了包含上述重组载体的重组菌株;所述菌株包括大肠杆菌或毕赤酵母;优选的,所述大肠杆菌为大肠杆菌DH5α,所述毕赤酵母为毕赤酵母GS115。
另一方面,本发明还提供了一种制备重组葡萄糖氧化酶的方法,所述方法包括,利用序列优化的葡萄糖氧化酶基因转化到毕赤酵母中,构建重组表达葡萄糖氧化酶的毕赤酵母,然后,对重组的毕赤酵母进行培养,并对重组的葡萄糖氧化酶进行纯化;优选的,所述毕赤酵母为毕赤酵母GS115。
另一方面,本发明还提供了固定化的葡萄糖氧化酶,所述葡萄糖氧化酶来源于上述方法重组表达的葡萄糖氧化酶。
另一方面,本发明还提供了上述密码子优化的葡萄糖氧化酶基因在制备葡萄糖氧化酶电极中的应用。
进一步的,所述葡萄糖氧化酶电极为将葡萄糖氧化酶固定在修饰的金电极上得到的葡萄糖氧化酶电极。
进一步的,所述修饰的金电极为采用半胱胺和纳米金修饰的金电极。
另一方面,本发明还提供了上述密码子优化的葡萄糖氧化酶基因在制备葡萄糖生物传感器中的应用。
本发明根据编码同一氨基酸的不同的密码子在毕赤酵母中使用频率的差异,去除葡萄糖氧化酶原始基因中稀有密码子,避免反向重复序列的出现,还要保证稳定RNA二级结构,去除内含子属性的剪接位点,优化了葡萄糖氧化酶基因,优化后的序列在毕赤酵母中能够成功表达,纯化后的目的蛋白能够用于制备葡萄糖氧化酶电极。
附图说明
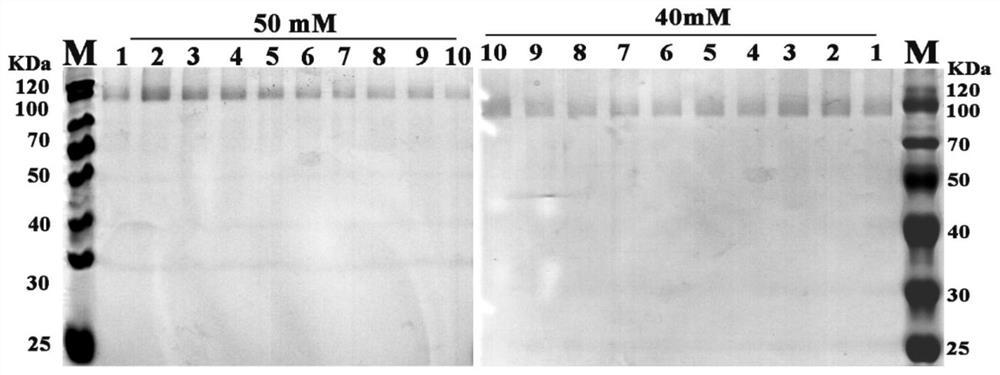
图1.在毕赤酵母中重组表达、纯化密码子优化的葡萄糖氧化酶,利用不同浓度咪唑洗脱重组蛋白电泳图,左图为利用50mM的咪唑洗脱效果图,右图为利用40mM的咪唑洗脱效果图,泳道1-10为特定浓度咪唑洗脱重组蛋白时每1mL为1收集管依次获得的密码子优化的葡萄糖氧化酶洗脱液。
图2.在毕赤酵母中重组表达、纯化未经密码子优化的葡萄糖氧化酶,利用不同浓度咪唑洗脱重组蛋白电泳图,左图为利用50mM的咪唑洗脱效果图,右图为利用40mM的咪唑洗脱效果图,泳道1-10为特定浓度咪唑洗脱重组蛋白时每1mL为1收集管依次获得的未经密码子优化的葡萄糖氧化酶洗脱液。
图3.重组葡萄糖氧化酶电极电化学表征图。
实施方式
下面结合实施例对本发明做进一步的说明,以下所述,仅是对本发明的较佳实施例而已,并非对本发明做其他形式的限制,任何熟悉本专业的技术人员可能利用上述揭示的技术内容加以变更为同等变化的等效实施例。凡是未脱离本发明方案内容,依据本发明的技术实质对以下实施例所做的任何简单修改或等同变化,均落在本发明的保护范围内。
实施例1、葡萄糖氧化酶基因密码子优化及克隆
本实施例中以AspergillusnigerAn76基因组中葡萄糖氧化酶g9669.t1基因为原始基因,原始基因序列如SEQ ID No.1所示。根据编码同一氨基酸的不同的密码子在毕赤酵母中使用频率的差异,去除葡萄糖氧化酶g9669.t1原始基因中稀有密码子,避免反向重复序列的出现,还要保证稳定RNA二级结构,去除内含子属性的剪接位点,优化了葡萄糖氧化酶基因,根据不同的策略,优化了两条序列,分别如SEQ ID No.2和SEQ ID No.3所示。
合成优化后的葡萄糖氧化酶基因,根据同源重组的原理利用南京诺唯赞 Ultra One Step Cloning Kit产品将密码子优化的葡萄糖氧化酶基因同源重组到pPIC9K质粒载体上,反应体系为10μL(1μL利用EcoRI和NotI线性化的pPIC9K载体,2μL葡萄糖氧化酶基因PCR片段,5μL2×ClonExpress Mix,2μL ddH2O),吸打混匀后,50℃反应15min,立即置于冰上冷却,即得葡萄糖氧化酶基因与pPIC9K质粒载体同源重组产物。
实施例2、葡萄糖氧化酶基因转化大肠杆菌富集质粒
分别将原始的葡萄糖氧化酶基因(SEQ ID No.1)以及密码子优化的葡萄糖氧化酶基因(SEQ ID No.2、SEQ ID No.3)与pPIC9K同源重组产物与E.coli DH5α混合,热激90s后涂布于100ug/mL氨苄抗性的LB琼脂培养平板上,37℃过夜培养。挑取单菌落,然后提取质粒电泳检测,并在-20℃保存质粒。再利用EcoRI与NotI酶切检测目的片段,之后送菌悬液由公司进行测序,将测序正确的质粒以同样方法转化大肠杆菌实现质粒富集。
实施例3、葡萄糖氧化酶基因转化毕赤酵母宿主菌
接种毕赤酵母GS115单菌落到含有5mLYPD液体培养基的试管中,30℃过夜培养。按1%接种量转接到含有50mL YPD液体培养基的三角瓶,30℃培养过夜,直到OD600=1.3~1.5;1500g,4℃条件下离心培养液5min,弃上清液,利用50mL冰浴双蒸水重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,利用25mL冰浴双蒸水重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,用2mL冰浴的1M的山梨醇溶液重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,用1mL冰浴的1M的山梨醇溶液重悬细胞,使菌悬液体积大约为1.5mL;将80μL处理好的感受态细胞和5~20μg经过SacI线性化了的实施例1-2得到的重组了葡萄糖氧化酶的质粒加入一个1.5mL预冷离心管中,混匀。然后把混合液转移入预先冰浴的转化杯中(0.2cm型);冰浴装有转化混合液的转化杯5min;按照Biorad毕赤酵母电转参数设置电转化仪(Voltage(V):2000;Capacitance(μF):25;Resistance(Ω):200;Cuvette(mm):2),并启动电脉冲,脉冲后立即往转化杯中加入1mL冰浴的1M的山梨醇溶液,然后把转化液转入一个新的1.5mL离心管中;30℃静置培养2h。吸取GS115转化液100μL涂布MD平板,30℃培养,直到转化子出现。对转化的MD平板上的单克隆进行菌落PCR验证,确保外源基因的整合。
实施例4、葡萄糖氧化酶基因诱导表达
接种筛选出来的毕赤酵母重组子于5mL BMGY液体培养基中,30℃,250rpm,振荡培养过夜;取500μL过夜培养物转接于50mL BMGY液体培养基中,30℃,250rpm,振荡培养至OD600=2~6(对数生长期,大约16-18h);3000g,离心5min,弃上清,用BMMY液体培养基重悬细胞至OD600=1.0,甲醇终浓度为1%;4)置于500mL三角瓶中,用8层灭菌纱布封口,30℃,250rpm,振荡培养;SDS-PAGE检测有无外源基因的表达;结果显示,原始的葡萄糖氧化酶基因(SEQ ID No.1)在毕赤酵母中无法成功表达,密码子优化后的葡萄糖氧化酶基因(SEQ IDNo.2)在毕赤酵母中能够成功表达,但是,SEQ ID No.3所示的密码子优化的葡萄糖氧化酶基因在毕赤酵母中也无法成功表达;这可能是因为此种密码子优化方法获得的葡萄糖氧化酶基因在毕赤酵母表达中受除密码子之外的因素影响导致其无法表达,尽管可以根据密码子偏好性对原始基因序列进行优化,但是,这只是理论上存在可能,在实际操作时能否在毕赤酵母中顺利表达,会与多种因素密切相关。
实施例5、密码子优化的葡萄糖氧化酶的分离纯化
将SDS-PAGE检测到目的蛋白表达的粗酶液与含有镍的填料混合,于4℃冰箱中转动结合6h。然后用浓度为10mM和20mM的咪唑溶液洗脱杂蛋白,用40mM和50mM咪唑溶液洗脱目的葡萄糖氧化酶,SDS-PAGE检测目的蛋白(图1)。用pH 5.0的磷酸氢二钠-柠檬酸缓冲液超滤40mM和50mM咪唑洗脱下来的蛋白溶液,4900rpm,4℃,直至流下来的缓冲液的pH为5.0,停止超滤,收集超滤获得的酶溶液。图1示出了利用密码子优化后的葡萄糖氧化酶基因(SEQID No.2)在毕赤酵母中成功表达后,利用不同浓度咪唑洗脱的重组蛋白的电泳图,由图1可知,采用40mM和50mM咪唑洗脱,能得到纯化的重组葡萄糖氧化酶。图2示出了采用原始的葡萄糖氧化酶基因(SEQ ID No.1)在毕赤酵母表达的纯化图谱,由于该原始的基因无法顺利表达,因此,在采用40mM和50mM咪唑洗脱时,无法纯化得到目的蛋白;SEQ ID No.3所示的密码子优化的葡萄糖氧化酶基因,由于无法成功表达,也没有纯化得到目的蛋白。
实施例6、密码子优化的葡萄糖氧化酶固定化
预处理裸金电极:打磨金电极先将金电极(φ=1mm)用去离子水洗净,再依次用φ=0.05μm及50μm的Al203抛光粉进行抛光处理直至镜面,用蒸馏水淋洗后进行超声清洗1min,取出自然晾干,再用硝酸(V:V=1:1),乙醇(V:V=1:1)依次超声清洗1min,取出用蒸馏水淋洗电极表面。组装前将电极放入0.5mol/L的硫酸中于-0.4—1.5V范围内循环扫描至稳定,取出用蒸馏水淋洗电极表面,自然晾干备用。
铁氰化钾溶液中循环伏安扫描:称取0.0823g铁氰化钾和5.055g KNO3置于250mL烧杯中,加入蒸馏水搅拌使其溶解,再转移至250mL容量瓶定容摇匀,制成1.0×10-3mol/L的铁氰化钾溶液(含0.2mol/L KNO3),并在所配制的铁氰化钾溶液中利用循环伏安法对金电极进行扫描,电位差在80mV以内,并接近64mV。
半胱胺修饰金电极:称取0.3857g半胱胺溶于50mL烧杯中,制成浓度为0.1mol/L的半胱胺溶液,并将裸金电极在4℃下浸泡于该半胱胺溶液中过夜后,取出用磷酸盐缓冲溶液冲洗,自然晾干备用。
合成纳米金溶胶:基于柠檬酸盐还原金盐反应原理制备纳米金溶胶,先用移液管移取1%(wt)氯金酸1.00mL,向其中加入79.00mL的蒸馏水,制得溶液A。再分别取4.00mL1%柠檬酸钠溶液与0.05mL 1%(wt)的靴酸溶液混合,将这两种溶液混合的同时加入16.00mL的蒸馏水,制得溶液B。分别将A,B两种溶液加热至60℃,在剧烈搅拌的同时迅速将B溶液加入到A溶液中得到混合溶液,并将该混合溶液加热到沸腾状态持续10min,溶胶由浅黄色逐渐显现成无色而后直至酒红色再将溶胶取出至室温下冷却。用透射电子显微镜对所制取的纳米金溶胶的微观结构进行表征,观察该溶胶是否为纳米级别,该纳米金颗粒的粒径在80nm左右。
半胱胺-纳米金修饰金电极:在4℃的环境中,将半胱胺修饰的金电极置于纳米金溶胶中浸泡24h,在冰箱中保存备用。
葡萄糖氧化酶与交联剂交联:将实施例5纯化得到的葡萄糖氧化酶、EDC、NHS、过氧化氢酶溶于10.0mL pH 7.00的缓冲溶液中,得到含有葡萄糖氧化酶、EDC、NHS、过氧化氢酶,缓冲溶液的交联剂,并将制得的交联剂保存于冰箱中备用。
葡萄糖氧化酶的固定化:将半胱胺、纳米金修饰的金电极取出用蒸馏水清洗,晾干。浸泡于配制的交联剂中24h,之后用相应pH的缓冲溶液淋洗,制得的葡萄糖氧化酶电极即可保存备用。
实施例7、葡萄糖氧化酶电极电化学表征
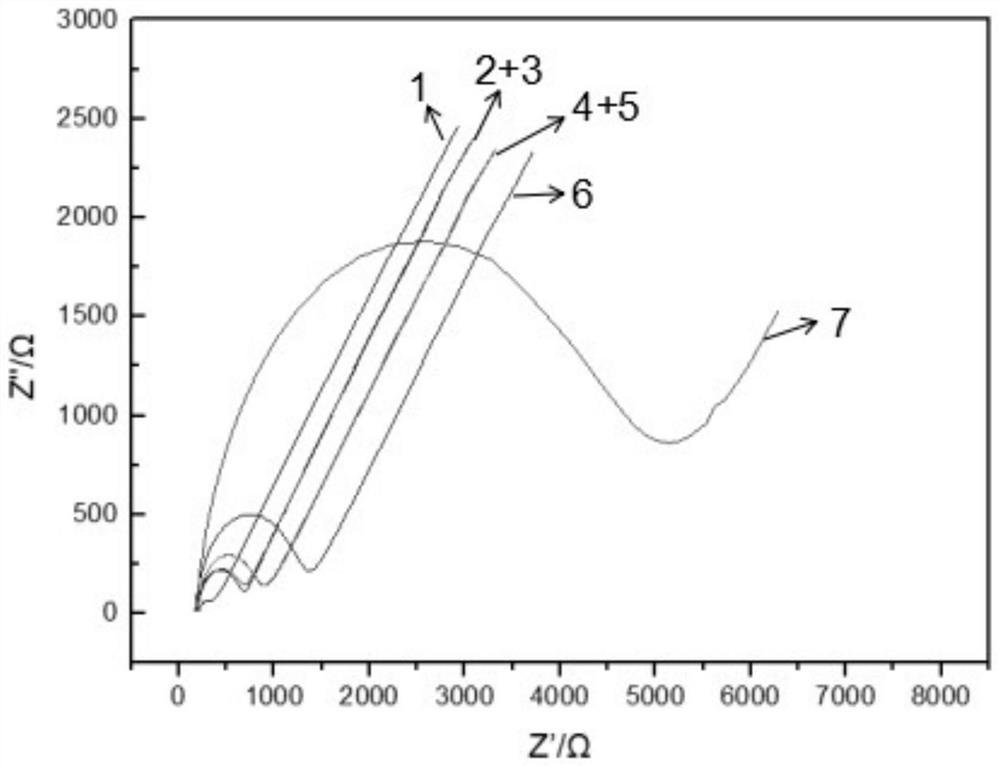
采用三电极体系对酶电极的电化学性质进行检测和探究,在电位范围为-0.4-1.6V时,依次对裸金电极、半胱胺修饰的金电极、纳米金-半胱胺修饰的电极及葡萄糖氧化酶-纳米金-半胱胺修饰的电极利用循环伏安法在的1.0×10-3mol/L铁氰化钾溶液(含0.2mol/L KNO3)中进行扫描检测,最后用葡萄糖氧化酶-纳米金-半胱胺修饰的电极对不同浓度的葡萄糖溶液(1mmol.L-1,1mmol.L-1,2mmol.L-1,4mmol.L-1,6mmol.L-1,8mmol.L-1,10mmol.L-1)进行循环伏安扫描,结果显示不同糖浓度条件下循环伏安曲线峰电位值与糖浓度呈良好的线性相关性(R2=0.996),如图3所示。
序列表
<110> 山东省科学院生物研究所
<120> 一种密码子优化的葡萄糖氧化酶基因及其应用
<160> 3
<170> SIPOSequenceListing 1.0
<210> 1
<211> 1788
<212> DNA
<213> 黑曲霉(Aspergillus niger)
<400> 1
ctcccacact acatcaggag caatggcatc gaagccagcc tcctgactga ccccaaggag 60
gttgccggcc gcactgtcga ctacatcatc gctggtggag gtctgactgg actcaccact 120
gctgcccgtc tgacggagaa ccccgatatc actgtgcttg tcatcgaaag tggctcctac 180
gagtctgaca gaggtcctat cattgaggac ctgaacgctt acggtgacat ttttggcagc 240
agtgtggacc acgcctacga gactgtcgag ctcgccacca acaatcagac tgcgctgatc 300
cgctccggaa atggtctcgg tggctctacc ctcgtcaacg gtggcacctg gactcgcccc 360
cacaaggcac aagttgactc atgggagacc gtcttcggaa atgagggctg gaactgggac 420
agcgtggccg cctactccct ccaggctgag cgtgctcgcg caccaaatgc caaacagatt 480
gctgctggcc actactttaa tgcatcctgc catggtatca atggtactgt ccacgccgga 540
ccccgcgata ccggtgatga ctactccccc atcgtcaagg ctctcatgag cgctgtcgaa 600
gacaggggcg ttcccaccaa gaaggacttg ggatgcggtg acccccatgg tgtgtccatg 660
ttccccaaca ccttgcacga agaccaagtg cgctctgatg ccgctcgcga atggctcctc 720
cccaactacc agcgtcccaa cctgcaagtc ctcactggac agtatgttgg aaaggtcctg 780
ctcagccaga acgctaccac acctcgtgcc gttggcgtgg aattcggcac ccacaagggc 840
aacacccaca acgtctacgc taagcacgag gtcctcctgg ccgctggatc cgctgtctct 900
cccaccatcc tcgaatattc cggtatcgga atgaagtcca ttctagagcc tcttggaatt 960
gacaccgtcg ttgacctgcc cgttggtctc aaccttcagg accagaccac ctctaccgtc 1020
cgctcacgca ttacctccgc cggtgccgga cagggacagg ccgcttggtt cgctaccttc 1080
aacgagacct ttggcgacta cgccgaaaag gctcacgagc tgctcaacac caagctggag 1140
cagtgggccg aagaggccgt cgcccgtggc ggattccaca acaccaccgc tttgctcatc 1200
cagtacgaga actaccgcga ctggatcgtc aaggacaatg tcgcatactc ggaactcttc 1260
ctcgacacgg ccggagtggc cagtttcgat gtgtgggatc ttctgccctt cactagagga 1320
tacgtacaca tcctcgacaa ggacccctac ctccgccatt tcgcatacga ccctcagtac 1380
tttctcaacg agcttgacct gctcggccag gctgccgcca ctcagctggc ccgcaacatc 1440
tccaactccg gtgccatgca aacttatttc gctggagaga ctattcccgg tgacaacctc 1500
gcgtatgatg ccgacttgag cgcctgggtt gagtatatcc cgtacaactt ccgccctaac 1560
taccatggtg tgggtacttg ctccatgatg ccgaaggaga tgggcggtgt tgtcgacaat 1620
gctgcccgtg tgtatggtgt gcagggactg cgagtcatcg atggttctat tccccctacg 1680
caaatgtcgt cccatgttat gacggtcttt tatgccatgg ccttgaagat tgcggatgcc 1740
atcttggcgg attacgcttc tatgcagcac caccaccacc accactaa 1788
<210> 2
<211> 1788
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 2
cttcctcact acatcagaag taatggaatt gaggctagtt tgttgaccga tccaaaggag 60
gttgctggta gaaccgtcga ctacatcatt gccggaggag gtcttactgg attgaccacc 120
gctgctaggt tgaccgagaa cccagacatc accgtccttg ttatcgagtc tggtagttac 180
gagagtgata gaggacctat catcgaggac ttgaacgcct atggtgacat ctttggttct 240
tctgtcgacc atgcctacga gaccgtcgaa ttggccacca ataaccagac cgccttgatt 300
agatctggaa atggattggg tggatctacc cttgtcaacg gtggtacttg gactagacca 360
cacaaggccc aagtcgactc ttgggagacc gtcttcggta atgaaggttg gaattgggac 420
agtgttgccg cttatagttt gcaagccgaa agggctagag cccctaatgc caaacagatc 480
gccgccggtc actatttcaa cgcttcttgc catggaatca acggtactgt ccacgctgga 540
ccaagagata ccggagacga ctattctcca attgtcaagg ccttgatgag tgctgtcgaa 600
gataggggtg ttccaaccaa aaaggacttg ggatgcggag atccacatgg tgtcagtatg 660
tttcctaata cccttcacga ggaccaagtt agaagtgatg ctgctagaga atggttgttg 720
cctaactacc agaggccaaa cttgcaagtc ttgaccggac agtacgtcgg taaggtcctt 780
ttgagtcaga acgccaccac tcctagagcc gtcggagttg agttcggaac tcacaagggt 840
aatacccaca acgtctacgc caaacacgag gtccttcttg ctgccggttc tgccgtctct 900
ccaaccatct tggaatactc tggaattgga atgaaatcta tcttggaacc tcttggtatc 960
gacaccgtcg ttgatttgcc agttggactt aaccttcaag atcagaccac cagtaccgtt 1020
aggagtagga tcacctctgc tggagctgga caaggacaag ctgcttggtt cgctaccttc 1080
aacgagacct tcggagacta tgccgagaag gcccacgagt tgcttaacac caagttggaa 1140
cagtgggccg aggaagctgt cgccagagga ggattccaca ataccaccgc cttgttgatt 1200
caatatgaaa actacagaga ctggatcgtc aaggacaacg tcgcttactc tgagttgttc 1260
cttgacactg ccggagttgc cagtttcgac gtctgggact tgcttccatt cactagaggt 1320
tatgtccaca tcttggataa agatccttac cttaggcact ttgcttatga tccacaatat 1380
tttcttaatg agttggactt gcttggtcaa gccgccgcta cccagttggc tagaaatatt 1440
agtaatagtg gtgccatgca aacctacttt gccggtgaga ccatccccgg agacaacttg 1500
gcttatgacg ccgacctttc tgcttgggtc gagtacattc cttacaactt tagaccaaac 1560
taccacggtg tcggtacttg tagtatgatg cctaaggaga tgggtggagt cgtcgataat 1620
gccgctaggg tttacggagt tcaaggactt agagttatcg acggttctat cccacctacc 1680
cagatgtctt ctcatgtcat gaccgtcttc tacgccatgg ctcttaagat cgctgacgcc 1740
atccttgctg actatgctag tatgcagcac caccaccatc atcattga 1788
<210> 3
<211> 1788
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
ctaccacatt atatccgatc taacggcatt gaagcttctc tattgactga tccaaaagag 60
gtcgctggaa ggactgtaga ttatattatc gctggcggtg gtttaactgg tttaaccacc 120
gctgctcgtt tgactgaaaa tcccgacatt actgtactag tgatcgagag tggaagttat 180
gaaagtgaca gaggtcccat cattgaagac ttgaatgctt acggtgacat cttcggttca 240
tcagttgatc atgcatacga gaccgttgaa ttggcaacaa acaaccaaac agctctaatt 300
cgttctggaa acggattggg tggttccacc ctggttaatg gtggtacatg gacgagacct 360
cacaaggcac aagttgattc atgggaaacc gtgttcggaa atgagggatg gaattgggac 420
agtgtagctg cctacagtct tcaagctgag agagctagag ccccaaacgc caagcagatc 480
gccgctggtc actattttaa tgcttcttgc cacggtatca acggcaccgt acacgctggt 540
ccaagagaca caggtgatga ttattctcct attgttaagg cattgatgtc cgcagttgaa 600
gacagaggtg ttcccacgaa gaaggaccta ggttgtggcg atcctcatgg cgtctctatg 660
tttcccaata ctttgcacga agatcaagta agatcagatg ccgcaaggga gtggctgctg 720
ccaaattacc aacgacctaa cttacaggtt cttacaggcc aatacgtggg taaagtgctg 780
ctttcccaga atgcaactac acccagggct gtaggtgtcg agttcggaac acataaggga 840
aatactcata acgtttacgc aaaacatgaa gtcttacttg ctgccggttc tgcagtatct 900
ccaactatcc tggaatattc tggcatcgga atgaagagta tccttgaacc actaggcata 960
gatacggtgg ttgatttgcc agtcggtcta aacctacaag atcagacaac ttccactgtc 1020
aggtccagaa tcacttctgc cggtgccgga caaggacaag cagcatggtt tgcaacattc 1080
aatgaaacct ttggtgacta cgccgagaag gctcacgagt tattgaacac aaagttagag 1140
caatgggcag aggaggccgt tgctcgtgga ggtttccata acacaaccgc tttgcttatt 1200
cagtacgaga attataggga ttggattgtt aaggataacg tcgcttattc agagttattc 1260
ctggatacgg ccggtgttgc atcattcgac gtatgggatt tgttgccttt tacaagaggt 1320
tacgtccaca tcttggacaa ggacccatat ctaagacatt ttgcatatga cccacaatat 1380
tttttgaacg aattggacct gcttggtcaa gccgctgcaa cccagttggc tcgaaatatt 1440
agtaactctg gcgctatgca aacctacttc gcaggagaaa cgataccagg agataatttg 1500
gcttacgatg ccgatttgtc tgcttgggtc gaatacatac catacaattt tcgtcctaat 1560
tatcacggtg tcggaacctg ttctatgatg cctaaggaaa tgggtggtgt agttgataac 1620
gctgctagag tatatggtgt ccagggattg cgtgttatcg acggaagtat tccacctaca 1680
cagatgtcat ctcatgttat gaccgtcttc tatgctatgg cactgaagat tgcagacgct 1740
atattggctg actacgcctc tatgcaacat caccatcacc atcattga 1788
一种密码子优化的葡萄糖氧化酶基因及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。
















动态评分
0.0