

IPC分类号 : C12N15/53I,C12N15/81I,C12N1/21I,C12N1/19I,C12N9/04I,C12Q1/00I,G01N27/48I,G01N27/327I,G01N27/30I,C12R1/84N,C12R1/19N




专利摘要
本发明提供了一种密码子优化的FAD‑葡萄糖脱氢酶基因,所述基因来源于AspergillusnigerAn76,其序列如SEQIDNo.2所示。该原始的FAD‑葡萄糖脱氢酶基因在毕赤酵母中无法成功表达,经过序列优化,其能够在毕赤酵母中顺利表达,并且,经过分离、纯化,能够得到纯化的目的蛋白,该重组FAD‑葡萄糖脱氢酶可以用于葡萄糖生物传感器中。
权利要求
1.一种密码子优化的FAD-葡萄糖脱氢酶基因,其特征在于,所述基因的核苷酸序列如SEQ ID No.2所示。
2.包含权利要求1所述基因的重组载体,所述载体骨架为pPIC9K载体。
3.包含权利要求2所述重组载体的重组菌株。
4.根据权利要求3所述的重组菌株,其特征在于,所述菌株包括大肠杆菌或毕赤酵母。
5.根据权利要求4所述的重组菌株,其特征在于,所述大肠杆菌为大肠杆菌DH5α。
6.根据权利要求4所述的重组菌株,其特征在于,所述毕赤酵母为毕赤酵母GS115。
7.一种制备重组FAD-葡萄糖脱氢酶的方法,其特征在于,所述方法包括如下步骤:
将权利要求1所述的基因转化到毕赤酵母中,构建重组表达FAD-葡萄糖脱氢酶的毕赤酵母,然后,对重组的毕赤酵母进行培养,并对重组的FAD-葡萄糖脱氢酶进行纯化;所述毕赤酵母为毕赤酵母GS115。
8.权利要求1所述的基因在制备FAD-葡萄糖脱氢酶电极中的应用。
9.根据权利要求8所述的应用,其特征在于,所述FAD-葡萄糖脱氢酶电极为将FAD-葡萄糖脱氢酶固定在修饰的玻碳电极上得到的葡萄糖脱氢酶电极。
10.根据权利要求9所示的应用,其特征在于,所述修饰的玻碳电极为采用二茂铁和多壁碳纳米管修饰的玻碳电极。
11.权利要求1所述的基因在制备葡萄糖生物传感器中的应用。
说明书
技术领域
本发明涉及生物酶基因工程和生物传感技术领域,具体涉及一种密码子优化的FAD-葡萄糖脱氢酶基因及其在制备生物传感元件中的应用。
背景技术
葡萄糖的准确、快速监测在医药、食品等行业有着重要意义,为此,生物传感器因其高特异性、分析时间短、可进行原位监测以及制造成本低等特点而成为葡萄糖监测的理想方法。虽然有这些优点,但是生物传感器仍然需要克服由用于检测的酶分子传感元件引起的问题。
在以酶为传感元件的生物传感器中,氧化还原酶是主要的生物识别元件,其中,葡萄糖氧化酶(GOX)是最常用的酶,GOX可以在氧气存在的条件下选择性的氧化葡萄糖成葡萄糖内酯,同时产生H2O2。然而,这类酶生物传感器的响应易受测量介质中氧浓度的影响,而使用人工电子介质会由于O2的存在导致葡萄糖检测信号的降低从而低估葡萄糖含量。此外,如果是基于测定H2O2水平反映葡萄糖含量,常常需要很高的工作电势(与标准电极相比,通常超过+600mv)对H2O2进行氧化,因此存在于生物性液体中的很多其他电活性化合物也可能被氧化而引起假电流响应。而基于烟酰胺腺嘌呤二核苷酸依赖性葡萄糖脱氢酶的生物传感器可以避免H2O2的产生,有望解决依赖H2O2检测葡萄糖的问题,但是这类酶必需向反应体系中额外添加昂贵的辅酶NAD+限制了这类酶的应用。因此,以吡咯喹啉醌(PQQ)或黄素腺嘌呤二核苷酸(FAD)为辅酶的葡萄糖脱氢酶(GDH)成为构建不以O2为电子受体或NAD+作为辅酶的生物传感器的合适候选酶类,但是部分GDH-PQQ分离自胞质外膜需要合适的洗涤剂溶解,而另外一部分水溶性GDH-PQQ特异性很低,这大大限制了这种酶类在生物传感器中的应用。GDH-FAD(FAD-葡萄糖脱氢酶基因)因其高催化效率、强底物专一性、低氧化还原电势以及不以O2作为电子受体的特点,使其成为构建不依赖O2的葡萄糖传感器最有应用潜力的酶分子元件。
真菌来源的GDH-FAD(EC 1.1.99.10)于1937年首次发现于米曲霉中,之后在土曲霉、黑曲霉、黄曲霉中也鉴定到GDH-FAD的存在。但是目前大部分GDH-FAD主要分离纯化自野生菌株,不容易进行重组表达,这也导致以GDH-FAD作为酶电极生物传感元件的研究较少,尤其是国内,只有少量关于GDH-FAD重组表达的研究,因此GDH-FAD高效重组表达及其在传感器领域的应用研究具有重要意义。
发明内容
本发明提供了一种密码子优化的FAD-葡萄糖脱氢酶基因。
本发明提供的密码子优化的FAD-葡萄糖脱氢酶基因来源于Aspergillus nigerAn76,其原始的基因序列如SEQ ID No.1所示,经过序列优化的基因序列如SEQ ID No.2所示。该原始的FAD-葡萄糖脱氢酶基因在毕赤酵母中无法成功表达,经过序列优化,其能够在毕赤酵母中顺利表达,并且,经过分离、纯化,能够得到纯化的目的蛋白。
另一方面,本发明还提供了包含FAD-葡萄糖脱氢酶基因的重组载体。本发明所述的载体包括克隆载体以及表达载体。
在一个实施方案中,所述克隆载体包括pUC系列的载体,例如,pUC18、pUC19;所述表达载体包括在大肠杆菌中表达的载体以及在毕赤酵母中表达的载体,优选的,包括pET系列的载体,如pET-21a;更优选的,所述在毕赤酵母中表达的载体为pPIC9K载体。
另一方面,本发明还提供了包含上述重组载体的重组菌株;所述菌株包括大肠杆菌或毕赤酵母;优选的,所述大肠杆菌为大肠杆菌DH5α,所述毕赤酵母为毕赤酵母GS115。
另一方面,本发明还提供了一种制备重组FAD-葡萄糖脱氢酶的方法,所述方法包括,利用序列优化的FAD-葡萄糖脱氢酶基因转化到毕赤酵母中,构建重组表达FAD-葡萄糖脱氢酶的毕赤酵母,然后,对重组的毕赤酵母进行培养,并对重组的FAD-葡萄糖脱氢酶进行纯化;优选的,所述毕赤酵母为毕赤酵母GS115。
另一方面,本发明还提供了固定化的FAD-葡萄糖脱氢酶,所述FAD-葡萄糖脱氢酶来源于上述方法重组表达的FAD-葡萄糖脱氢酶。
另一方面,本发明还提供了上述密码子优化的FAD-葡萄糖脱氢酶基因在制备FAD-葡萄糖脱氢酶电极中的应用。
进一步的,所述FAD-葡萄糖脱氢酶电极为将FAD-葡萄糖脱氢酶固定在修饰的玻碳电极上得到的FAD-葡萄糖脱氢酶电极。
进一步的,所述修饰的玻碳电极为采用二茂铁和多壁碳纳米管修饰的玻碳电极;优选的,所述二茂铁为羟甲基二茂铁,所述多壁碳纳米管为羧基化多壁碳纳米管
另一方面,本发明还提供了上述密码子优化的FAD-葡萄糖脱氢酶基因在制备葡萄糖生物传感器中的应用。
本发明根据编码同一氨基酸的不同的密码子在毕赤酵母中使用频率的差异,去除FAD-葡萄糖脱氢酶原始基因中稀有密码子,避免反向重复序列的出现,还要保证稳定RNA二级结构,去除内含子属性的剪接位点,优化了FAD-葡萄糖脱氢酶基因,优化后的序列在毕赤酵母中能够成功表达,纯化后的目的蛋白能够用于制备FAD-葡萄糖脱氢酶电极。
附图说明
图1.SDS-PAGE检测毕赤酵母GS115连续六天诱导表达的密码子优化的FAD-葡萄糖脱氢酶。M为蛋白标准品,1d、2d、3d、4d、5d、6d分别指诱导培养毕赤酵母GS115至1天、2天、3天、4天、5天、6天时取得的发酵粗酶液。
图2.SDS-PAGE检测利用不同浓度咪唑洗脱在毕赤酵母GS115中重组表达经密码子优化的FAD-葡萄糖脱氢酶,左图为利用10mM的咪唑洗脱效果图,右图为利用20mM的咪唑洗脱效果图,泳道1-10为特定浓度咪唑洗脱重组蛋白时每1mL为1收集管依次获得的经密码子优化的FAD-葡萄糖脱氢酶洗脱液。
图3.重组FAD-葡萄糖脱氢酶电极电化学表征图。
图4.重组FAD-葡萄糖脱氢酶电极检测不同浓度葡萄糖循环伏安响应曲线。
实施方式
下面结合实施例对本发明做进一步的说明,以下所述,仅是对本发明的较佳实施例而已,并非对本发明做其他形式的限制,任何熟悉本专业的技术人员可能利用上述揭示的技术内容加以变更为同等变化的等效实施例。凡是未脱离本发明方案内容,依据本发明的技术实质对以下实施例所做的任何简单修改或等同变化,均落在本发明的保护范围内。
实施例1、FAD-葡萄糖脱氢酶基因密码子优化及克隆
本实施例中以Aspergillus niger An76基因组中FAD-葡萄糖脱氢酶g5086.t1基因为原始基因,原始基因序列如SEQ ID No.1所示。根据编码同一氨基酸的不同的密码子在毕赤酵母中使用频率的差异,去除FAD-葡萄糖脱氢酶g5086.t1原始基因中稀有密码子,避免反向重复序列的出现,还要保证稳定RNA二级结构,去除内含子属性的剪接位点,优化了FAD-葡萄糖脱氢酶基因,根据不同的策略,优化了两条序列,分别如SEQ ID No.2和SEQ IDNo.3所示。
合成优化后的FAD-葡萄糖脱氢酶基因,根据同源重组的原理利用南京诺唯赞 Ultra One Step Cloning Kit产品将密码子优化的FAD-葡萄糖脱氢酶基因同源重组到pPIC9K质粒载体上,反应体系为10μL(1μL利用EcoRI和NotI线性化的pPIC9K载体,2μL FAD-葡萄糖脱氢酶基因PCR片段,5μL 2×ClonExpress Mix,2μL ddH2O),吸打混匀后,50℃反应15min,立即置于冰上冷却,即得FAD-葡萄糖脱氢酶基因与pPIC9K质粒载体同源重组产物。
实施例2、FAD-葡萄糖脱氢酶基因转化大肠杆菌富集质粒
分别将原始的FAD-葡萄糖脱氢酶基因(SEQ ID No.1)以及密码子优化的FAD-葡萄糖脱氢酶基因(SEQ ID No.2、SEQ ID No.3)与pPIC9K同源重组产物与E.coli DH5α混合,热激90s后涂布于100ug/mL氨苄抗性的LB琼脂培养平板上,37℃过夜培养。挑取单菌落,然后提取质粒电泳检测,并在-20℃保存质粒。再利用EcoRI与NotI酶切检测目的片段,之后送菌悬液由公司进行测序,将测序正确的质粒以同样方法转化大肠杆菌实现质粒富集。
实施例3、FAD-葡萄糖脱氢酶基因转化毕赤酵母宿主菌
接种毕赤酵母GS115单菌落到含有5mLYPD液体培养基的试管中,30℃过夜培养。按1%接种量转接到含有50mL YPD液体培养基的三角瓶,30℃培养过夜,直到OD600=1.3~1.5;1500g,4℃条件下离心培养液5min,弃上清液,利用50mL冰浴双蒸水重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,利用25mL冰浴双蒸水重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,用2mL冰浴的1M的山梨醇溶液重悬细胞;1500g,4℃条件下离心培养液5min,弃上清液,用1mL冰浴的1M的山梨醇溶液重悬细胞,使菌悬液体积大约为1.5mL;将80μL处理好的感受态细胞和5~20μg经过SacI线性化了的实施例1-2得到的重组了FAD-葡萄糖脱氢酶的质粒加入一个1.5mL预冷离心管中,混匀。然后把混合液转移入预先冰浴的转化杯中(0.2cm型);冰浴装有转化混合液的转化杯5min;按照Biorad毕赤酵母电转参数设置电转化仪(Voltage(V):2000;Capacitance(μF):25;Resistance(Ω):200;Cuvette(mm):2),并启动电脉冲,脉冲后立即往转化杯中加入1mL冰浴的1M的山梨醇溶液,然后把转化液转入一个新的1.5mL离心管中;30℃静置培养2h。吸取GS115转化液100μL涂布MD平板,30℃培养,直到转化子出现。对转化的MD平板上的单克隆进行菌落PCR验证,确保外源基因的整合。
实施例4、FAD-葡萄糖脱氢酶基因诱导表达
接种筛选出来的毕赤酵母重组子于5mL BMGY液体培养基中,30℃,250rpm,振荡培养过夜;取500μL过夜培养物转接于50mL BMGY液体培养基中,30℃,250rpm,振荡培养至OD600=2~6(对数生长期,大约16-18h);3000g,离心5min,弃上清,用BMMY液体培养基重悬细胞至OD600=1.0,甲醇终浓度为1%;4)置于500mL三角瓶中,用8层灭菌纱布封口,30℃,250rpm,连续振荡培养6天,每天取样1mL;SDS-PAGE检测有无外源基因的表达;结果显示,原始的FAD-葡萄糖脱氢酶基因(SEQ ID No.1)在毕赤酵母中无法成功表达,密码子优化后的FAD-葡萄糖脱氢酶基因(SEQ ID No.2)在毕赤酵母中能够成功表达(图1),但是,SEQ IDNo.3所示的密码子优化的FAD-葡萄糖脱氢酶基因在毕赤酵母中也无法成功表达;这可能是因为此种密码子优化方法获得的FAD-葡萄糖脱氢酶基因在毕赤酵母表达中受除密码子之外的因素影响导致其无法表达,尽管可以根据密码子偏好性对原始基因序列进行优化,但是,这只是理论上存在可能,在实际操作时能否在毕赤酵母中顺利表达,会与多种因素密切相关。
实施例5、密码子优化的FAD-葡萄糖脱氢酶的分离纯化
将SDS-PAGE检测到目的蛋白表达的粗酶液与含有镍的填料混合,于4℃冰箱中转动结合6h。然后用浓度为5mM的咪唑溶液洗脱杂蛋白,用10mM和20mM咪唑溶液洗脱目的FAD-葡萄糖脱氢酶,SDS-PAGE检测目的蛋白(图2)。用pH 5.0的磷酸氢二钠-柠檬酸缓冲液超滤10mM和20mM咪唑洗脱下来的蛋白溶液,4900rpm,4℃,直至流下来的缓冲液的pH为5.0,停止超滤,利用3K超滤管收集超滤获得的酶溶液。图2示出了利用密码子优化后的FAD-葡萄糖脱氢酶基因(SEQ ID No.2)在毕赤酵母中成功表达后,利用不同浓度咪唑洗脱的重组蛋白的电泳图,由图2可知,采用10mM和20mM咪唑洗脱,能得到纯化的重组FAD-葡萄糖脱氢酶(0.48mg/mL)。
实施例6、密码子优化的FAD-葡萄糖脱氢酶固定化
预处理玻碳电极:打磨玻碳电极先将玻碳电极用去离子水洗净,再依次用φ=0.05μm及50μm的Al2O3抛光粉进行抛光处理直至镜面,用蒸馏水淋洗后进行超声清洗1min,取出自然晾干,再用硝酸(V:V=1:1),乙醇(V:V=1:1)依次超声清洗1min,取出用蒸馏水淋洗电极表面,自然晾干备用。
铁氰化钾溶液中循环伏安扫描:称取0.0329g铁氰化钾和2.022g KNO3置于烧杯中,加入80mL蒸馏水搅拌使其溶解,再转移至100mL容量瓶定容摇匀,制成1.0×10-3mol/L的铁氰化钾溶液(含0.2mol/L KNO3),并在所配制的铁氰化钾溶液中利用循环伏安法对玻碳电极进行扫描,电位差在80mV以内,并接近64mV。
羧基化多壁碳纳米管修饰玻碳电极:称取0.3g羧基化多壁碳纳米管置于溶于50mL烧杯中,加入0.5g盐酸一乙基一3-(3一二甲基氨基丙基)碳二亚胺(EDC)和0.5g N-羟基琥珀酰亚胺(NHS),并将三者溶解于10mL蒸馏水中,室温静置活化碳管6h。再用13000×g离心10min,弃上清取出沉淀碳管,再用适量蒸馏水复溶,重复该步离心并加蒸馏水清洗,直至将碳管洗至中性,再烘干备用。称取上述制备的碳管0.3g超声溶于100mL蒸馏水中,制成浓度为3g/L的羧基化多壁碳纳米管溶液,取7uL羧基化多壁碳纳米管滴涂在玻碳电极表面,自然晾干备用。
羟甲基二茂铁-多壁碳纳米管修饰玻碳电极:在4℃的环境中,将羧基化多壁碳纳米管修饰的玻碳电极置于1g/L的羟甲基二茂铁溶液中浸泡24h,在冰箱中保存备用。
FAD-葡萄糖脱氢酶与交联剂交联:称取0.5g壳聚糖溶于100mL蒸馏水中,边搅拌边滴加醋酸直至壳聚糖全部溶解,制成0.5%的壳聚糖溶液。将实施例5纯化得到的FAD-葡萄糖脱氢酶(1mg/mL)、0.5%壳聚糖溶液、25%戊二醛溶液按体积等比例混合,得到含有FAD-葡萄糖脱氢酶、壳聚糖、戊二醛的交联酶溶液,并将制得的交联酶溶液保存于4℃中备用。
FAD-葡萄糖脱氢酶的固定化:将羟甲基二茂铁-多壁碳纳米管修饰的玻碳电极取出用蒸馏水清洗,晾干。滴加7μL配制的交联酶溶液在4℃下晾干,之后用pH 7.0的磷酸缓冲溶液淋洗,即制得FAD-葡萄糖脱氢酶电极,4℃保存备用。
实施例7、FAD-葡萄糖脱氢酶电极电化学表征
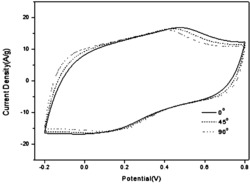
采用三电极体系对酶电极的电化学性质进行检测和探究,在电位范围为-0.8-0.8V时,依次对羟甲基二茂铁-多壁碳纳米管修饰的玻碳电极在双蒸水、1mM Glucose中以及FAD-葡萄糖脱氢酶-壳聚糖-羟甲基二茂铁-多壁碳纳米管修饰的玻碳电极在1mMGlucose中利用循环伏安法进行扫描检测,结果显示,在玻碳电极只修饰羟甲基二茂铁-多壁碳纳米管而未修饰FAD-葡萄糖脱氢酶时,无论是在水中还是葡萄糖溶液中都没有氧化还原峰,而修饰了FAD-葡萄糖脱氢酶以后在葡萄糖溶液中检测时,在电位为+0.4和+0.2处出现明显的氧化还原峰,说明FAD-葡萄糖脱氢酶电极可以实现葡萄糖的检测,最后用FAD-葡萄糖脱氢酶-壳聚糖-羟甲基二茂铁-多壁碳纳米管修饰的玻碳电极对不同浓度的葡萄糖溶液(3.8mmol.L-1,5.8mmol.L-1,6.3mmol.L-1,6.9mmol.L-1,7.2mmol.L-1,8.0mmol.L-1)进行循环伏安扫描,结果如图4所示,随着糖浓度的增加氧化峰对应的电流值逐渐增大,说明制得的FAD-葡萄糖脱氢酶电极可以作为传感元件应用到葡萄糖生物传感器中,实现样品中葡萄糖的定量检测。
序列表
<120> 一种密码子优化的FAD-葡萄糖脱氢酶基因及其应用
<160> 3
<170> SIPOSequenceListing 1.0
<210> 1
<211> 1722
<212> DNA
<213> 黑曲霉(Aspergillus niger)
<400> 1
ggctcacccg ctcactatga ttttgtcatt gtcggtggag gcaccagtgg attggttgtt 60
gccaacagac tctcggaact gaataatgtc actgtagccg tcattgaggc cggtgaatct 120
gcattgaaca acttcaatgt gtctaatgtc atgggctaca gcacggcatt cgggacggag 180
gtcgactggg cctaccaaac cgagaaccag acatatgcag gaggcttgca gcagactatc 240
cgtgccggaa aggcacttgg aggtacaagc acaatcaatg gaatgtccta cactcgggcc 300
gaagacgtgc aaattgacaa ttgggaagtg ctcgggaacg aaggttggaa ctggaagaac 360
ctgtttcaat attataagaa gtcggaaggc ttccaggtgc cgaccaagga ccagatcgct 420
catggtgcca actacaacgc tagctatcat ggtttgaacg gccctctgaa ggttggctgg 480
cctacctcca tgaccaacag cagcgtcttc cctgtccttg aacagacctt cgagaaactg 540
ggcgttcagt acaaccccga ttccgaaggc ggcaagatgg tcggattcac tgttcacccc 600
gacactctgg accgagagat gaatgttcgc gaagatgctg ctagggctta ctactggcca 660
tatgaagccc gctcgaacct gaagatcatt tctaatactc gtgcaaacaa ggtcatctgg 720
gccgatacca cccagggaga ggccgttgcc gtcggcattg aagttactaa cgcttacggc 780
acgcaaacca tctacgctga caaggaggtc atcctgtcgg ccggcgcgct tagatcccct 840
gccctgcttg agctgtctgg tatcggaaac cccgacgtcc tcaataagca caatatcccc 900
gtcaaggtca acatcaccac cgtcggggag aacctacagg atcagaccaa caacgctctt 960
tcctgggaag gcgtcgacac cctcactggt ttggcaacct tctccgtcct gccatctgtg 1020
aaccagctct acggcgataa tgtcactgcc ttggcttcct acgtcaagtc ccaacttgct 1080
agctatgcta agaccgttgc tgatgcctcc aatggcgctg tgaaggaggc caatctcatt 1140
gaggctttcg agcgccagta cgacctgatc ttcaactccc aggtccccta tgcagaagtc 1200
gttttcgctc cgagcggaca atcgttcgcg gtcgaatact ggcctcttct tcccttctcc 1260
cgcggcagcg ttcatatcca gtctgcgaat gcttcagatt accctgccat caatcccaac 1320
tacttcatgt tcagacagga tgccgatgct caaatcacgg tggcacagta cattcggaag 1380
gctttgggaa ctacacctct gaacagtctt gtgggcgagg aagtctcacc cagtcttgaa 1440
gtgctccctg cgaatgcgtc cagctccact tggactaagt gggttcaaga aaactaccga 1500
accaactacc accctgttgg caccaacagc atgcttcccc gcgaaaaggg tggtgttgtc 1560
agccccgagc tcagggttta cggtaccaag aatcttcgcg tcgtcgatgc atcagtcctg 1620
ccattccagc tctgcggcca tctgaccagc actctgtatg ctgtcgctga aagggcttcc 1680
gatctgatta aggaaagtta ccaccaccac caccaccact aa 1722
<210> 2
<211> 1722
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 2
ggttctccag ctcattacga ttttgttatt gttggaggtg gtactagtgg tttggttgtt 60
gctaatagat tgtctgaatt gaataacgtt accgttgctg ttattgaggc tggtgagtct 120
gctttgaata attttaatgt ttccaacgtc atgggttact ctactgcttt tggtactgaa 180
gtcgattggg cttatcagac tgaaaatcaa acttatgctg gaggtcttca acaaactatt 240
agagccggta aggctttggg tggtacttct actattaacg gaatgtctta tactagagct 300
gaggatgttc aaattgataa ctgggaagtt ttgggtaatg aaggttggaa ctggaagaac 360
ttgtttcaat attataagaa gtccgaggga tttcaagttc ctactaagga ccaaattgct 420
catggtgcta actacaacgc tagttaccat ggtttgaacg gtccattgaa ggtcggatgg 480
cctacttcca tgaccaactc ttcagttttt ccagttcttg aacaaacctt cgaaaagttg 540
ggtgttcaat acaacccaga ttctgaagga ggtaagatgg ttggtttcac tgttcatcca 600
gacactttgg acagagaaat gaatgttaga gaggatgcag ctagagctta ctactggcca 660
tatgaagcta gatccaactt gaagattatt tcaaacacta gagctaacaa ggtcatttgg 720
gccgatacta cccaaggtga ggctgttgct gttggtattg aagttactaa cgcttacggt 780
acacaaacca tctacgctga taaggaagtt atcttgtcag ctggagcatt gagatctcct 840
gctttgttgg aattgtctgg tattggtaat ccagatgttt tgaacaagca taacattcca 900
gttaaggtca acattactac tgttggtgaa aacttgcagg atcaaactaa caacgctttg 960
tcttgggaag gtgttgatac cttgaccggt cttgctactt tttctgtttt gccatctgtt 1020
aaccaattgt acggagataa tgttactgct ttggcttctt atgttaagtc ccaattggct 1080
tcttacgcta agactgttgc tgatgcttct aacggtgctg ttaaggaggc taacttgatt 1140
gaggcttttg aaagacaata tgatttgatt ttcaactccc aagttccata cgctgaggtt 1200
gtttttgccc catctggtca atcttttgca gttgaatatt ggccattgtt gccattttct 1260
agaggttctg ttcatattca atctgctaac gcttctgatt atccagccat taatccaaat 1320
tactttatgt ttagacagga cgctgatgct caaattactg ttgctcagta cattagaaag 1380
gctttgggaa ctactccact taattctttg gttggtgaag aagtttcccc atccttggaa 1440
gttttgcctg ccaacgcttc ttcttctacc tggaccaagt gggttcaaga aaactacaga 1500
actaactacc acccagttgg tactaactct atgttgccaa gagaaaaagg tggtgttgtt 1560
tcccctgaat tgagagttta cggtactaag aacttgagag ttgtcgatgc ctctgttttg 1620
ccttttcaac tttgtggtca cttgacttct accctttacg ctgtcgctga aagagcttcc 1680
gatttgatta aggaatctta ccatcatcat catcaccatt aa 1722
<210> 3
<211> 1722
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
ggaagtcctg cccactacga ttttgtgatt gttggtggtg gcacttccgg acttgtcgtt 60
gctaatagat tatccgaatt gaacaatgtc acagtggccg ttatagaagc cggtgagtcc 120
gctcttaaca actttaatgt gtcaaatgta atgggctact ctactgcttt cggaactgaa 180
gtggattggg cataccaaac agaaaaccaa acttacgccg gtggtctaca acagactatt 240
cgtgctggaa aggccttggg aggaacctca actatcaatg gtatgtctta tactagagct 300
gaggacgtgc aaattgataa ttgggaagtc ttgggaaatg aaggatggaa ctggaaaaac 360
ttattccagt actacaaaaa atctgaagga tttcaggtac ctaccaagga tcagattgct 420
cacggtgcca actataacgc ttcatatcat ggactaaatg gtccattgaa ggtcggttgg 480
cctaccagta tgactaactc aagtgtgttc ccagttctag aacaaacatt cgaaaaactt 540
ggagtgcagt acaacccaga cagtgaagga ggtaaaatgg ttggattcac tgtccatcct 600
gacactttgg atcgtgagat gaatgtcaga gaggatgccg caagagccta ttactggcct 660
tacgaagcta gatcaaattt aaaaattatt tccaacacga gagctaacaa ggtcatctgg 720
gcagatacga cccaaggaga ggctgtggct gtcggtattg aagtcacaaa tgcttatgga 780
acacaaacta tatatgctga caaagaagtt atcctgtccg caggtgcttt gcgatctcca 840
gccttgttgg aattatccgg cattggtaat cctgacgttt taaacaagca caatatccct 900
gttaaggtga acataactac cgttggcgag aatttgcaag atcagactaa caacgcactt 960
tcatgggagg gtgttgatac attgactgga ttggcaacat tcagtgtact accctctgtt 1020
aatcaattgt acggtgataa tgttactgcc ctggcctcat atgtgaaatc tcaattggca 1080
tcatatgcta aaacggtagc cgacgcaagt aacggtgccg taaaagaagc taaccttatc 1140
gaagctttcg agagacaata cgatttaatc tttaacagtc aggttccata tgcagaggtc 1200
gtcttcgcac catcaggtca gtcattcgct gttgaatatt ggcccttatt gccattttca 1260
agaggaagtg tccacattca atctgcaaac gcctccgatt accctgctat aaaccctaat 1320
tactttatgt tccgtcagga tgcagacgct caaattactg ttgcccaata tataagaaaa 1380
gctctaggta cgacgccttt aaactcttta gttggtgagg aagtctcacc atctctggaa 1440
gttctgcctg caaacgctag ttcatctacc tggacaaagt gggtccagga gaactacaga 1500
accaactacc atccagtggg aaccaattct atgctgccaa gagaaaaagg aggagtagtc 1560
tcccctgagc ttcgtgtcta tggtaccaaa aacttgagag ttgttgatgc atctgtcttg 1620
ccattccagc tgtgtggcca ccttacctct actttgtacg ctgttgccga gagagcttca 1680
gacttgatca aggaatccta tcatcaccac caccatcatt aa 1722
一种密码子优化的FAD-葡萄糖脱氢酶基因及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。

















动态评分
0.0