

IPC分类号 : C12N9/12,C12N9/02,C12N15/54,C12N15/53,C12N15/63,C12N5/10,C12N1/00,C12N15/11,C12N15/82







专利摘要
本发明公开了一种植物耐旱相关蛋白及其编码基因与应用。本发明提供的蛋白质,是如下(a)或(b)的蛋白质:(a)由序列表中序列2所示的氨基酸序列组成的蛋白质;(b)将序列2的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与植物耐旱性相关的由序列2衍生的蛋白质。本发明还保护编码所述蛋白的基因。所述基因可用于培育耐旱植物。本发明的耐旱相关蛋白及其编码基因为人为控制植物中耐旱相关基因的表达提供了基础,将在培育耐旱植物中发挥重要的作用。
说明书
技术领域技术领域
本发明涉及一种植物耐旱相关蛋白P5CS及其编码基因与应用。
技术背景背景技术
棉花是我国重要的纤维作物,在国民经济中占有重要地位。但是我国棉花种植区大部分处于干旱(如新疆棉区)和半干旱地区(如黄河流域棉区),目前干旱已经成为棉花生产的主要限制因素之一。
在干旱和盐胁迫时,植物通过渗透调节防止水分的过度散失,保持细胞的膨压,从而保持细胞的生长、气孔开放和光合作用等生理过程的正常进行。在所有的有机渗透调节物质中,脯氨酸是分布最广的渗透调节剂。它主要以游离状态广泛存在于植物中。作为植物体内一种重要的渗透调节物质,脯氨酸主要有渗透调节、保护生物大分子以及参与氮代谢和能量代谢等作用。近年来已通过转基因技术使许多植物产生更多的渗透调节保护物质脯氨酸,从而提高了植物的耐旱性。吡咯琳-5-羧酸合成酶(Pyrroline-5-carboxyate synthetase,P5CS)是脯氨酸合成途径中的关键酶。
发明内容发明内容
本发明的目的是提供一种植物耐旱相关蛋白P5CS及其编码基因与应用。
本发明提供的植物耐旱相关蛋白(GaP5CS),来源于亚洲棉(Gossypiumarboreum),是如下(a)或(b)的蛋白质:
(a)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(b)将序列2的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与植物耐旱性相关的由序列2衍生的蛋白质。
序列表中的序列2由716个氨基酸残基组成,包含一个谷氨酸激酶结构域(Glutamyl kinase,GK)(自序列2的氨基末端第14至259位氨基酸残基)和一个谷氨酸-γ-半醛脱氢酶结构域(Glu-5-semialdehyde dehydrogensae,GSA)(自序列2的氨基末端第286至691位氨基酸残基)。
所述一个或几个氨基酸残基的取代和/或缺失和/或添加是指将序列2的上述两个结构域外的氨基酸残基进行取代和/或缺失和/或添加。
为了使(a)中的GaP5CS便于纯化,可在由序列表中序列2所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。
表1标签的序列
上述(b)中的GaP5CS可人工合成,也可先合成其编码基因,再进行生物表达得到。上述(b)中的GaP5CS的编码基因可通过将序列表中序列1自5′端第351至2498位核苷酸所示的DNA序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。
编码上述植物耐旱相关蛋白的基因(GaP5CS)也属于本发明的保护范围。
所述基因可为如下1)或2)或3)或4)的DNA分子:
1)其编码序列是序列表中序列1自5’第351至2498位核苷酸所示的DNA分子;
2)序列表中序列1所示的DNA分子;
3)在严格条件下与1)或2)限定的DNA序列杂交且编码所述蛋白的DNA分子;
4)与1)或2)或3)限定的DNA序列具有90%以上同源性,且编码耐旱相关蛋白的DNA分子。
上述严格条件可为在6×SSC,0.5%SDS的溶液中,在65℃下杂交,然后用2×SSC,0.1%SDS和1×SSC,0.1%SDS各洗膜一次。
序列表中的序列1所示由2827个核苷酸组成,自5’第1至350位核苷酸为5’非编码区(5’-UTR)(350bp),第351至2498位核苷酸为编码序列(2148bp),第2499至2501位核苷酸为终止密码子,第2502至2827位核苷酸为3’非编码区(3’-UTR)(326bp)。
含有以上任一所述基因的重组表达载体也属于本发明的保护范围。
可用现有的植物表达载体构建含有所述基因的重组表达载体。
所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。所述植物表达载体还可包含外源基因的3’端非翻译区域,即包含聚腺苷酸信号和任何其它参与mRNA加工或基因表达的DNA片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mRNA前体的3’端,如农杆菌冠瘿瘤诱导(Ti)质粒基因(如胭脂合成酶Nos基因)、植物基因(如大豆贮存蛋白基因)3’端转录的非翻译区均具有类似功能。
使用所述基因构建重组植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型启动子或组成型启动子,如花椰菜花叶病毒(CAMV)35S启动子、玉米的泛素启动子(Ubiquitin),它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是ATG起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(GUS基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
所述重组表达载体可将所述基因插入pBI21质粒的多克隆位点得到。具体来说,所述重组表达载体可为pBI121/GaP5CS;所述pBI121/GaP5CS是将pBI21质粒BamH I和Sac I位点间的小片段取代为所述基因(GaP5CS)得到的。
含有以上任一所述基因(GaP5CS)的表达盒、转基因细胞系及重组菌均属于本发明的保护范围。
扩增所述基因(GaP5CS)全长或任一片段的引物对也属于本发明的保护范围。
本发明的另一个目的是提供一种培育耐旱转基因植物的方法。
本发明所提供的培育耐旱转基因植物的方法,可将编码所述植物耐旱相关蛋白的基因导入目的植物(如植物细胞或组织)中,得到耐旱能力高于目的植物的转基因植物。具体来说,可以将所述重组表达载体导入目的植物中,得到耐旱能力高于目的植物的转基因植物。
本发明同时提供了一种培育持水力提高的转基因植物的方法,是将所述植物耐旱相关蛋白的基因导入目的植物(如植物细胞或组织)中,得到持水力高于目的植物的转基因植物。具体来说,可以将所述重组表达载体导入目的植物中,得到持水力高于目的植物的转基因植物。作物离体叶片在空气中的失水速率(单位时间的失水量)反映叶片的持水力,或称植物组织抗脱水能力。离体叶片失水速率与植株的抗旱性之间有密切关系。
利用任何一种可以引导外源基因在植物中表达的载体,将编码所述蛋白的基因导入植物细胞,可获得耐旱能力增强的转基因细胞系及转基因植株。携带有所述基因的表达载体可通过使用Ti质粒、Ri质粒、植物病毒载体、直接DNA转化、显微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物组织培育成植株。被转化的植物宿主既可以是单子叶植物,也可以是双子叶植物,如:烟草、百脉根、拟南芥、水稻、小麦、玉米、黄瓜、番茄、杨树、草坪草、苜宿等。
本发明发现了一种植物耐旱相关蛋白(GaP5CS)及其编码基因(GaP5CS)。GaP5CS基因可被PEG6000诱导表达。将转化GaP5CS基因后得到的过量表达的烟草植株进行逆境生理实验,证明转入GaP5CS基因后,提高了烟草的耐旱性。本发明的耐旱相关蛋白及其编码基因为人为控制植物中耐旱相关基因的表达提供了基础,将在培育耐旱植物中发挥重要的作用。
以下结合附图及具体实施例进一步阐述本发明。
附图说明附图说明
图1为GaP5CS基因受到PEG6000胁迫条件诱导表达。
图2为pBI121-GaP5CS表达载体的构建示意图。
图3为获得的抗卡那霉素转pBI121/GaP5CS烟草;ck:对照烟草;1-2:转基因烟草。
图4为转pBI121/GaP5CS烟草基因组PCR检测结果;M:Marker;CK:对照烟草;1、2:转基因烟草。
图5为干旱条件下转pBI121/GaP5CS烟草和对照烟草比较;CK:对照烟草;1、2:转基因烟草。
图6为转基因烟草叶片在脱水过程中失水速率的变化;CK:对照烟草;L1、L2:转基因烟草。
图7为转基因烟草叶片在脱水过程中离子渗漏的变化;CK:对照烟草;L1、L2:转基因烟草。
具体实施方式具体实施方式
以下的实施例便于更好地理解本发明,但并不限定本发明。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂商店购买得到的。
棉花(石系亚1号):购自中国农业科学院棉花研究所棉花种质资源中期库,库编号:I3A01616。
实施例1、植物耐旱相关蛋白(GaP5CS)及其编码基因的发现
提取3-6片真叶幼苗期棉花(石系亚1号)幼苗总RNA,反转录得其全基因组的cDNA。将cDNA作为模板,进行RT-PCR扩增。
上游引物:5’-ATGGCTGAGGATAGTTCAAGAGCTTTTGTT-3’;
下游引物:5’-TCAAGAATTTATTGGTTGTCTTTGTGGGTGT-3’。
PCR扩增体系(50ul):MgCl2(25mM)2ul,10×Buffer 5ul,dNTP Mixture(10uM)4ul,Easy-A High-Fidelity PCR Cloning Enzyme(5u/ul)1ul,反转录反应液5ul(0.25ug),上游引物(10uM)2ul,下游引物(10uM)2ul,ddH2O补齐至50ul。PCR条件:94℃预变性3min;94℃30s,55℃40s,72℃5min,30个循环;72℃延伸10min。
反应结束后,将得到PCR产物进行测序鉴定。在测序结果的基础上,通过3’和5’RACE技术(Takara)获得PCR产物上游和下游的核苷酸序列。
最终获得序列表中序列1所示的核苷酸序列,该核苷酸序列具有完整的编码框。将序列1所示的核苷酸序列命名为GaP5CS。
GaP5CS基因的核苷酸序列如序列表中序列1所示,由2827个核苷酸组成。序列表的序列1自5’第1至350位核苷酸为5’非编码区(5’-UTR)(350bp),第351至2498位核苷酸为编码序列(2148bp),第2499至2501位核苷酸为终止密码子,第2502至2827位核苷酸为3’非编码区(3’-UTR)(326bp)。
GaP5CS基因编码序列表的序列2所示的蛋白质(GaP5CS)。GaP5CS由716个氨基酸组成,预测分子量为77.5kD,等电点(pI)7.019。
TargetP软件预测GaP5CS基因表达产物为细胞质定位;无跨膜螺旋;蛋白质功能域预测发现GaP5CS包含两个功能域:谷氨酸激酶结构域(Glutamyl kinase,GK)和谷氨酸-γ-半醛脱氢酶(Glu-5-semialdehyde dehydrogensae,GSA)。因此,GaP5CS基因能编码一个催化合成脯氨酸合成的酶,从而在棉花耐旱中起到功能。
实施例2、植物耐旱相关蛋白(GaP5CS)的诱导表达
将石系亚1号幼苗在28℃恒温培养箱中培养至3-6片真叶,小心拔出幼苗进行干旱胁迫处理。17%PEG6000(质量百分含量)处理0、0.5、1、2、3、4、5、6h后,分别提取幼苗总RNA(总RNA纯度与质量都较高,每个样品RNA的A260/A280都在1.8-2.1之间,浓度在1.0ug/ul以上)。分别将不同处理时间的幼苗的总RNA进行1%琼脂糖凝胶电泳检测,28SrRNA与18SrRNA条带非常清晰,表明所提取的RNA样品能用于RT-PCR反应。
将不同处理时间的幼苗的总RNA进行RT-PCR分析,以棉花看家基因Histone 3(GENBANK ACCESSION NUMBER:AF024716)为内参,作为衡量模板量的对照。
RT-PCR引物如下:
扩增内参引物:
上游引物:5’-TCAAGACTGATTTGCGTTTCCA-3’;
下游引物:5’-GCGCAAAGGTTGGTGTCTTC-3’;
扩增GaP5CS基因的引物:
上游引物:5’-TGAAAGGAAGTGGGCAGGTAGT-3’;
下游引物:5’-TTCTACTCAAATTCTCCTGACACA-3’。
结果如图1所示。PEG6000模拟干旱胁迫处理时,GaP5CS基因的mRNA迅速积累并在胁迫2h时达到最大值,PEG胁迫4h后,GaP5CS的转录本开始下降,之后在胁迫6h时又开始回升。
实施例3、耐旱转基因植株的获得与耐旱性鉴定
一、重组表达载体的构建
构建流程见图2。
1、制备带有BamH I和Sac I酶切位点的GaP5CS基因(编码区;序列表的序列1自5’第351至2498位核苷酸)。
2、用BamH I和Sac I双酶切植物表达载体pBI121(购自Clontech;Cat.#6018-1);用BamH I和Sac I双酶切步骤1制备的GaP5CS基因;将酶切后的载体骨架和酶切后的GaP5CS基因连接,得到重组质粒。
3、将得到的重组质粒进行测序,测序结果表明:GaP5CS基因插入到了pBI21的35S启动子下游,将携带GaP5CS的pBI121重组载体命名为:pBI121/GaP5CS。
二、耐旱转基因植株的获得
将步骤一构建的pBI121/GaP5CS,冻融法转化至农杆菌LBA4404(购自Invitrogen;Cat.No.18313-015),采用叶盘转化法转化野生型烟草NC89(购自中国农业科学院烟草研究所烟草种质资源中期库,库编号:I5A01846);将pBI121冻融法转化至农杆菌LBA4404(购自Invitrogen;Cat.No.18313-015),采用叶盘转化法转化野生型烟草NC89,得到转空载体对照植株;具体步骤如下:
1、烟草种子用2.5%次氯酸钠处理8min,用无菌水洗一次,然后用70%乙醇浸泡1min,无菌水洗3~4次,播种于MS培养基上,于25℃,16h光照/8h黑暗条件培养5周。
2、将烟草叶片剪去边缘和叶脉,切成0.4cm×0.6cm大小;切好的烟草叶盘用重组农杆菌菌液浸泡3-5min;用无菌滤纸吸干植物材料表面的菌液,转入烟草共培养基中,25℃黑暗条件下共培养两天;将转化处理的烟草叶盘浸于MS液体培养基(含500mg/L羧苄青霉素)中,浸泡3~5min后用无菌滤纸吸去残留液,转入分化培养基(50-100mg/L浓度的卡纳霉素)中,约一周继代一次,直到分化出幼芽;将幼芽切下,置入生根培养基中诱导生根。获得的抗卡那霉素的转基因烟草(T0代)见图3。
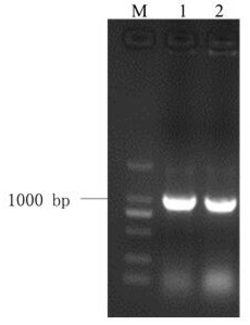
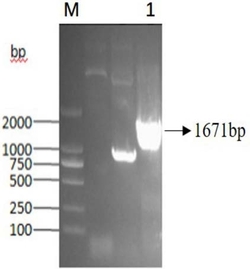
3、提取抗卡那霉素转pBI121/GaP5CS烟草和转空载体对照植株的基因组DNA,进行PCR检测,烟草中可能含有目的基因的同源基因,为保证PCR检测的特异性,根据载体启动子序列和外源转化基因的保守区设计引物,来区别外源基因和内源基因。引物为:5’-GCTCCTACAAATGCCATCA-3’;5’-CTGCACATGCCTTCCCATCAAGTT-3’。
PCR检测结果见图4。结果表明得到了含有目的基因的转基因植株。
三、T0代转基因烟草耐旱性测定
烟草为盆栽土培,正常供水,定期施MS营养液。以转空载体的烟草作为转基因烟草的对照(每个株系50棵)。均设置三次重复试验,结果取平均值。
1、自然干旱处理
在棉花所温室内,相同条件培育转基因烟草、对照烟草和野生型烟草NC89,待植株生长至4个月左右时,停止供水5天后观察植株的萎蔫程度。对照植株和野生型烟草NC89明显萎蔫,转基因植株受影响程度明显好于对照烟草和野生型烟草NC89,对照烟草和野生型烟草NC89表型一致。转pBI121/GaP5CS烟草和对照烟草比较见图5。
2、离体叶片失水速率测定
作物离体叶片在空气中的失水速率(单位时间的失水量)反映叶片的持水力,或称植物组织抗脱水能力。离体叶片失水速率与植株的抗旱性之间有密切关系,且与某些产量和植株性状存在较明确的关系。检测转基因植株、对照植株和野生植株离体叶片的失水速率,方法如下:
相同条件培育转基因烟草、对照烟草和野生型烟草NC89,待植株生长至4个月时,快速剪取生长一致的烟草新鲜叶片,迅速装入塑料袋密封蔽光。在温湿度相对稳定的实验室内称取初始叶重后,空气中放置9h自然风干后称重,最后于105℃烘干30min,80℃烘干10h,测干重,计算单位重量的叶片在单位时间的失水量。离体叶片失水速率由RWL=(IW-Wi)/{DW×(Ti-T0)}计算。式中IW为T0时初始叶重(g),Wi为Ti时叶重(g),DW为干重(g),T0和Ti分别为称取IW和某Wi的时间(h)。
在空气中自然风干9h后,烟草转基因株系的叶片失水速率明显小于对照烟草和野生型烟草NC89,对照烟草和野生型烟草NC89的叶片失水速率一致,说明GaP5CS的转入增强了烟草的持水力。转基因烟草和对照烟草叶片在脱水过程中失水速率的变化结果见图6。
3、叶片细胞的质膜透性测定
干旱胁迫会导致细胞膜结构的破坏,叶片离子渗漏反映细胞质膜的完整性,检测转基因植株、对照植株和野生植株叶片的电导率,方法如下:
相同条件培育转基因烟草、对照烟草和野生型烟草NC89,待植株生长至4个月时,快速剪取生长一致的烟草新鲜叶片,迅速装入塑料袋密封蔽光。在温湿度相对稳定的实验室内称取初始叶重后,空气中放置9h自然风干。风干后的烟草叶片放入75ml去离子水中(25℃),2h后测定电阻率R1,测完R1的样本100℃煮沸20分钟,冷却至25℃测定电阻率R2。离子渗漏=(R1/R2)×100%。
在空气中自然干燥9h后,烟草转基因株系的叶片离子渗漏小于对照烟草和野生型烟草NC89,对照烟草和野生型烟草NC89的叶片离子渗漏一致,表明干旱胁迫条件下,转基因植株对干旱逆境膜伤害的抵抗能力显著增强,推测这与脯氨酸所具有的对生物膜结构的保护作用有关。转基因烟草和对照烟草叶片在脱水过程中离子渗漏的变化结果见图7。
一种植物耐旱相关蛋白P5CS及其编码基因与应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。










动态评分
0.0