

IPC分类号 : C07K7/08,C07K14/00,C12N15/11,A61K38/10,A61K38/16,A61P35/00









专利摘要
本发明提供了一种非共价靶向CRM1的多肽及其应用,该多肽的氨基酸序列选自SEQIDNO:1‑SEQIDNO:10任一项所示的氨基酸序列。本发明提供的非共价靶向CRM1的多肽,约10个氨基酸,能与CRM1以非共价方式结合,该结合方式可逆,且毒副作用很小,对肿瘤有很好的抑制作用,且不容易产生耐药性,为广谱抗肿瘤药物提供理论基础。
权利要求
1.非共价靶向CRM1的多肽,其特征在于,所述多肽的氨基酸序列如SEQ ID NO:10所示。
2.一种DNA片段,其特征在于,包含编码权利要求1所述多肽的核苷酸序列。
3.一种表达载体,其特征在于,包含权利要求2所述的DNA片段。
4.一种宿主细胞,其特征在于,包含权利要求2所述的DNA片段或权利要求3所述的表达载体。
5.权利要求1所述的多肽或权利要求2所述的DNA片段在制备肿瘤抑制剂中的应用。
6.一种药物组合物,其特征在于,包括权利要求1所述的多肽或权利要求2所述的DNA片段以及药学上可接受的赋形剂。
说明书
技术领域
本发明属于生物医药技术领域,具体涉及一种非共价靶向CRM1的多肽及其应用。
背景技术
细胞核是真核细胞内最大、最重要的细胞结构,是细胞遗传与代谢的调控中心。核被膜作为细胞核与细胞质的界膜,它通过其上的核孔复合物(NPC)调控着细胞核内外的物质交换与信息交流。NPC是一个双功能,双向性的亲水性核质运输通道。也就是其不仅可以介导蛋白质的入核转运,也介导RNA、核糖核蛋白颗粒(RNP)、蛋白质等的出核运输。而这些分子量大于近40KD的功能性蛋白质的出入核都必须要先通过与转运因子形成复合物,然后由转运因子介导着一起参与核质运输。
CRM1由20个热重复结构(Heat Repeat)组成,是非常重要的出核转运因子,其运输机制(如图1所示):CRM1在细胞核内可以识别含有NES出核信号肽的蛋白质cargo,然后在核内RanGTP的协同促进作用下形成三元复合物出核,运往细胞质后,该CRM1-RanGTP-NES·cargo三元复合物在RanGAP和RanBP1的作用下去组装,CRM1卸货,cargo去参与其下游的信号通路,维持生物功能。
现有文献报道,CRM1在多种肿瘤细胞中高表达。CRM1的高表达,导致了细胞核内的许多含有NES的功能蛋白(如p53、p21、APC等)被CRM1过度错误定位到细胞质里,扰乱了细胞的正常功能,导致肿瘤等多种疾病。所以,针对出核因子CRM1识别NES·cargo的分子机制,基于结构靶向性设计药物,抑制CRM1的运输通路,可以有效杀死肿瘤细胞。
现已有的第一代、第二代抑制剂如LMB、KPT-SINE等,是一系列通过与CRM1的NES槽上的528位半胱氨酸共价不可逆结合的小分子,虽然杀伤力强,但存在以下缺陷:
(1)LMB与CRM1-C528形成的共价键是不可逆的,同时SINE-CRM1共价键的水解也非常缓慢。因为CRM1是人体所必须的出核转运因子,不可逆的LMB和水解非常缓慢的SINE会持久抑制CRM1,导致其在临床测试上具有较大毒性而不利后期临床的推进。
(2)LMB、SINE等小分子的反应基团在体内和体外能够和很多不同的小分子作用,尤其是SINE可以结合任何含有半胱氨酸的多肽。所以给药测试时可能会导致一些非特异性结合,引起一些不必要的副作用,降低药物有效性。
(3)由于LMB,SINE等依赖结合CRM1-528位上的半胱氨酸(Cys528)来抑制核内蛋白质出核。病人如果产生Cys528突变,或者本来存在此突变,将对共价结合在Cys528的突变体产生明显的抗药性。
发明内容
针对现有技术中存在的上述问题,本发明提供一种非共价靶向CRM1的多肽及其应用,可有效解决现有技术中存在的与CRM1共价结合的小分子具有不可逆,毒副作用大,有效性差,不耐受528位半胱氨酸突变的问题。
本发明的目的之一是基于CRM1与NES作用的分子机制,靶向结构,优化设计非共价靶向CRM1的多肽,设计的非共价靶向CRM1的多肽及氨基酸序列如下:
Nm2:GSTVEDMTKKFGTLTIHDT(SEQ ID NO:1);
Nm3:GSTVDELTKKFGTLTIHDT(SEQ ID NO:2);
Nm4:GSTVEDLTKKFGTLTIHDT(SEQ ID NO:3);
Nm12:GSTVDEMTKKFGTLTIHDDD(SEQ ID NO:4)
Nm13:GSDDTVDEMTKKFGTLTIHDT(SEQ ID NO:5);
Nm15:GSDDTVDELTKKFGTLTIHDDD(SEQ ID NO:6);
Nm16:GSDDTVEDMTKKFGTLTIHDDD(SEQ ID NO:7);
Nm17:GSDDTVEDLTKKFGTLTIHDDD(SEQ ID NO:8);或
Nm42:DGYIEELIRMFGKLSIHDD(SEQ ID NO:10)。
本发明的第二个目的是提供一种DNA片段,该DNA片段包含编码上述多肽的核苷酸序列。
本发明的第三个目的是提供一种表达载体,该表达载体包含上述的DNA片段。
本发明的第四个目的是提供一种宿主细胞,该宿主细胞包含上述DNA片段。
本发明的第五个目的是将上述多肽或DNA片段用于制备肿瘤抑制剂中的应用。
本发明的第六个目的是提供一种药物组合物,该药物组合物包括上述所述的多肽或DNA片段以及药学上可接受的赋形剂。
本发明提供的非共价靶向CRM1的多肽及其应用,具有以下有益效果:
本发明提供的非共价靶向CRM1的多肽,约10个氨基酸,能与CRM1以非共价方式结合,该结合方式可逆,且毒副作用很小,对肿瘤有很好的抑制作用,且不容易产生耐药性,为广谱抗肿瘤药物提供理论基础。
附图说明
图1为CRM1的核转运机制。
图2为MVM和CRM1/Ran/RanBP1的晶体结构图。
图3为MVM与CRM1的疏水槽的相互作用图。
图4为PD检测分析各NES突变体与CRM1的结合强弱结果图。
图5为ITC定量分析各NES突变体与CRM1的亲和力结果图。
图6为Nm2与CRM1的晶体结构。
图7为Nm12与CRM1的晶体结构。
图8为Nm13与CRM1的晶体结构。
图9为Nm15与CRM1的晶体结构。
图10为GFP-NES与CRM1在293T,HeLa细胞中的共定位分析结果。
图11为GFP-NES免疫共沉淀293T,HeLa细胞裂解中的CRM1量分析结果。
图12为Nm15的核输出抑制作用结果图。
图13为细胞存活率与NES出核率分析结果图。
图14为Nm42与CRM1亲和力分析结果图。
图15为Nm42与CRM1的结构。
图16为iDPP/DNA纳米复合物模拟图。
图17为iDPP/Nm42纳米复合物对肿瘤细胞B16-F10 cells的存活率分析结果图。
图18为小鼠活体成像结果图。
图19为iDPP/Nm42在体内的抗肿瘤活性结果图。
图20为iDPP/Nm42的安全评估结果图。
图21为iDPP/Nm42对小鼠体重的影响结果图。
图22为iDPP/Nm42对小鼠ALT、AST和CREA水平的影响结果图。
图23为KPT-330和Nm42对CRM1的C528S突变细胞的耐药性分析结果图。
具体实施方式
实施例1解析NS2 NES与CRM1/Ran/RanBP1的晶体结构
NS2是小鼠微小病毒(MVM)的一个结构蛋白,它上面有一段超生理的NES,与CRM1的亲和力较高,以致可以在没有RanGTP的协同促进条件下与CRM1的NES疏水槽稳定结合。所以,首先基于这段NS2 NES(后面简称MVM)与CRM1结合的分子机制来设计优化一些亲和力更高的非共价结合CRM1的多肽。
为了从结构上观察MVM与CRM1的相互作用,本发明解析了MVM/CRM1/Ran/RanBP1四元复合物的晶体结构。选择四元复合物的原因(1)CRM1/Ran/RanBP1三元复合物的晶体结构已有报导,为本发明四元复合物的结构解析奠定了基础。(2)本发明选择的四元复合物中的CRM1的H9 loop(406-426)是删掉的,以致于在结构上该CRM1的NES结合槽是开放的,有利MVM的结合。最终形成稳定的MVM/CRM1/Ran/RanBP1四元复合物。具体方案如下:
(1)MVM的克隆,表达纯化:
先将MVM的序列(GSTVDEMTKKFGTLTIHDT)克隆到pGEX-4T1的表达载体上的GST标签后面,中间用TEV酶切位点相连,然后将重组好的质粒转化到BL21(DE3)表达宿主中,用1升LB培养基培养。在OD为0.8~1.0时,加入0.5mM IPTG诱导剂,18℃诱导15个小时。之后将大肠杆菌菌液4000rpm离心,弃上清,用20mL悬菌buffer(50mM Tris pH 8.0,200mM NaCl,10%glycerol,2mM DTT,1mM EDTA和1mM PMSF)重悬。接着将菌液进行600-900Mpa高压破碎,15000rpm离心40min,将上清与GST-beads柱在4℃旋转结合2个小时,之后将GST-beads用wash buffer(20mM Tris8.0 200mM NaCl,10%Glycerol,2mM EDTA,2mM DTT)洗6次,每次14mL。最后用20mM Tris pH8.0,200mMNaCl,1mM EDTA,2mM DTT,10mM GSH buffer将GST-MVM洗脱下来。浓缩至1mL,用Superdex 200进行纯化。取峰值蛋白跑12%SDS-PAGE,检查蛋白的纯度。并将纯度较高的蛋白分装,并测蛋白浓度,储存在-80℃以供实验所需。
(2)CRM1,Ran,RanBP1的表达纯化:
CRM1与Ran的蛋白均用pET-28a表达,用载体上面的His标签进行纯化。CRM1用TB培养基,在OD约1.8-2.2时,加入IPTG,在25℃过夜表达10个小时。而Ran用LB培养基表达,在37℃诱导。之后收菌时,由于CRM1与Ran的纯化是用Ni-beads纯化,所以其悬菌buffer用50mMTris pH8.0,10mM Imidazole pH 7.5,300mM NaCl,2mM MgCl2,10%甘油,1mM PMSF。在清洗beads上的杂蛋白时,所用wash buffer是20mM Tris pH8.0,10mM Imidazole pH 7.5,300mM NaCl,2mM MgCl2,10%甘油。最后Ni beads的洗脱buffer是300mM Imidazole pH7.5,300mM NaCl,2mM MgCl2,10%甘油,2mM BME。纯化的方式与前面一样。
最后RanBP1的表达纯化与GST-MVM一样。不同的是本发明会用TEV酶将GST标签从RanBP1上面切掉,所用方法是将TEV蛋白酶与纯化的GST-RanBP1蛋白进行4℃过夜孵育,然后将混合液蛋白进行阳离子柱纯化。最终得到纯的RanBP1蛋白。
(3)四元复合物的获得与结晶:
将以上纯化好的蛋白(CRM1,RanBP1,RanGTP,GST-MVM)按一定比例混合,保证CRM1少量,RanGTP稍过量,GST-MVM完全过量,同时体积应小于1mL,此时它们已形成有四元复合物。然后加入少量TEV酶,4℃过夜酶切掉GST-MVM,保证四元复合物没有GST标签。然后用buffer(10mM Tris 7.5,100mM NaCl,5mM Mg(OAc)2,0.1mM GTP,2mM BME)经S200i纯化,分离。最终取峰值两边蛋白跑12%SDS-PAGE,检查复合物的形成。取形成复合物的峰,浓缩至6mg/mL,在16℃环境中,在0.1M Bis-Tris(pH 6.6),0.2M NH4NO3和18%PEG3350的条件下进行悬滴结晶。
(4)晶体的获得与结构的解析:
当晶体长到一定程度,以最快的速度在显微镜下将晶体捞出,并过一遍防冻buffer(结晶条件再补加12%甘油),立即放入液氮并送上海光源(SSRF)收集X-光衍射数据。之后利用已有的yCRM1-hRan-yRanBP1(pdb code:4HAT)结构模型来搭建解析并优化本发明的结构数据。
实施例2基于MVM-CRM1的结构设计NES突变体
由以上方案,本发明解析到了MVM的结构。发现MVM符合class 1a NES类型,与PKI结合CRM1的类型相同(如图2和图3所示,图2为MVM和CRM1/Ran/RanBP1的晶体结构图,图3为MVM与CRM1的疏水槽的相互作用图)。从结构上可以看到D80/E81
若将MVM突变成E80/D81,其与CRM1的亲和力有可能会上升,所以本发明设计了Nm2:GSTVEDMTKKFGTLTIHDT(SEQ ID NO:1)。将NES序列上Φ1位置的氨基酸突变成L(亮氨酸)亲和力会更高,所以本发明设计了Nm3:GSTVDELTKKFGTLTIHDT(SEQ ID NO:2)。本发明也将Nm2和Nm3进行了共同突变,设计了Nm4:GSTVEDLTKKFGTLTIHDT(SEQ ID NO:3)。在结构上观察,发现MVM的两端都特别靠近CRM1的碱性电子云表面,所以推测把MVM的两端分别加上两个酸性氨基酸DD(天冬氨酸),可能会增加其与CRM1的亲和力,所以本发明设计了Nm12:GSTVDEMTKKFGTLTIHDDD(SEQ ID NO:4)、Nm13:GSDDTVDEMTKKFGTLTIHDT(SEQ ID NO:5)。
本发明把Nm2-4与Nm12,Nm13突变到一起,也分别设计了Nm15-17,Nm15:GSDDTVDELTKKFGTLTIHDDD(SEQ ID NO:6);Nm16:GSDDTVEDMTKKFGTLTIHDDD(SEQ ID NO:7);Nm17:GSDDTVEDLTKKFGTLTIHDDD(SEQ ID NO:8)。
突变体和WT的序列汇总如下:
WT(MVM):GSTVDEMTKKFGTLTIHDT(SEQ ID NO:9);
Nm2:GSTVEDMTKKFGTLTIHDT(SEQ ID NO:1);
Nm3:GSTVDELTKKFGTLTIHDT(SEQ ID NO:2);
Nm4:GSTVEDLTKKFGTLTIHDT(SEQ ID NO:3);
Nm12:GSTVDEMTKKFGTLTIHDDD(SEQ ID NO:4)
Nm13:GSDDTVDEMTKKFGTLTIHDT(SEQ ID NO:5);
Nm15:GSDDTVDELTKKFGTLTIHDDD(SEQ ID NO:6);
Nm16:GSDDTVEDMTKKFGTLTIHDDD(SEQ ID NO:7);
Nm17:GSDDTVEDLTKKFGTLTIHDDD(SEQ ID NO:8)。
实施例3检测NES突变体与CRM1的亲和力
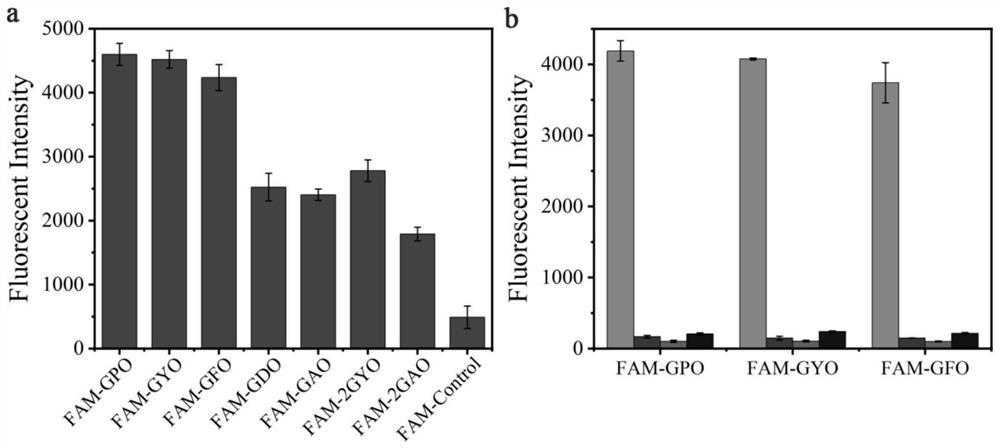
Pull-down(PD)检测法:首先本发明在体外构建表达了GST-MVM,GST-NES突变体,并通过分子筛S200i纯化。为了测试这些突变体与CRM1的相互作用,将一定量的GST标签的MVM及突变体与GSH beads孵育,同时加入定量的CRM1可溶性蛋白,用PD buffer定容到1ml后共同在4℃环境下孵育结合1.5个小时,然后用PD buffer(20mM Tris pH 7.5,200mMNaCl,10%glycerol,2mM MgCl2,0.005%Triton-X100和2mM DTT)清洗3次,洗掉没有结合到beads上的蛋白,最后将结合到beads上的蛋白通过SDS/PAGE跑胶和考马斯亮蓝观察。每个实验至少重复2遍。通过PD检测,筛选出Nm15与CRM1的亲和力最高(如图4所示)。
Isothermal Titration Calorimetry(ITC)检测法:在20℃温度下,用ITC200(Microcal)仪器,在20mM Tris pH 8.0,200mM NaCl and 2mM MgCl2 buffer条件下。用125μM GST-NES突变体分别滴定相同的复合物(包括12μM yRanBP1,8μM hCRM1,和10μMRanGTP)。每个实验重复至少两次,同时用NITPIC和SEDPHAT分析ITC数据定量分析NES亲和力的变化。通过ITC定量分析,筛选出的Nm15与CRM1的亲和力相比MVM提升了近10倍,亲和力在nM级别(如图5所示)。
实施例4解析几个亲和力较高的NES突变体与CRM1复合物的晶体结构
采用解析NS2 NES与CRM1/Ran/RanBP1的晶体结构方法来获得并解析Nm2(如图6所示),Nm12(如图7所示),Nm13(如图8所示)和Nm15(如图9所示)的晶体结构。
图6中将MVM和Nm2与CRM1的相互作用进行了对比分析,绿色的NES是MVM,银色的是Nm2。可以看到将MVM的D80/E81突变成Nm2的E80/D81后,Nm2的这两个氨基酸分别与CRM1上面的K533/N571存在相互作用。
图7中紫色标记的Nm12的末端一个D(天冬氨酸)与CRM1的R542存在电荷相互作用。
图8中黄色标记的Nm13的末端一个D(天冬氨酸)与CRM1的K525存在电荷相互作用。
图9中蓝色表示的Nm15中其优化的三处氨基酸突变82L(异亮氨酸),和两端均突变加上的D(天冬氨酸)同时与CRM1相互作用。
所获的上述结构分辨率在 具有较好的电子云密度,数据可信度较高。可以看到在NM15上,M82L的突变及两端额外的相互作用在结构上似乎与CRM1契合得更好,它们在提高NES与CRM1的亲和力上可能提供着重要的贡献。总之,这些突变体在结构上的分析进一步验证了本发明的猜想,也进一步符合了本发明在PD和ITC上的数据结果。
实施例5细胞上观察最优多肽Nm15与CRM1的亲和力
基于体外NES的设计优化,接下来构建了pEGFP-Nm15和pEGFP-WT哺乳动物细胞表达质粒。接着在体内比较了GFP-Nm15与GFP-WT的细胞定位和与CRM1的胞内结合情况。
细胞共定位实验:在293T和Hela细胞中分别转染一定量的GFP-Nm15、GFP-WT和GFP,培养一段时候后,将细胞进行固定,染色处理,在confocal显微镜下观察,成像,统计发现GFP-Nm15与CRM1有一个特别强的共定位(如图10所示)。由图10可知,GFP-Nm15在细胞内的共定位比GFP-WT和GFP强很多。
Immunoprecipitate(IP)实验:将上诉转染有GFP-Nm15、GFP-WT和GFP的细胞进行裂解,并用GFP或IgG抗体beads进行免疫共沉淀,之后通过SDS/PAGE,转膜,CRM1和GFP抗体孵育,接着相应物种二抗孵育后,最后用显色液曝光观察,结果如图11所示。由图11可知,在Input同样量的情况下,GFP-Nm15 IP下细胞裂解液中CRM1的量最多,由此可知,在细胞内,Nm15与CRM1的亲和力最高。
实施例6细胞内观察Nm15对CRM1核输出和细胞生长的抑制作用
核输出抑制作用:在293T和Hela细胞中,首先转染mCherry-NES-MBP-NLS出核蛋白(图中标记为NES),接着再分别用H2O、KPT-330或者转染GFP-N1,GFP-WT、GFP-Nm15质粒进行处理。培养24小时后,细胞被处理,固定,并用Hoechst染色。最后用Olympus FV-1000confocal显微镜进行观察,成像,并用NIH ImageJ软件进行分析统计,结果见图12。图12-1是HeLa和A549细胞经不同处理后的细胞Confocal成像分析图,可以看到相比与WT,在加入GFP-Nm15后,NES货物基本被抑制到核内无法出核。图12-2是对细胞核内NES量的统计数据,结果一致。因此,Nm15与WT相比,在细胞内对CRM1的核输出抑制作用更强。
细胞生长抑制作用:直接对293T和Hela细胞分别进行DMSO、KPT-330或者GFP-N1,GFP-WT、GFP-Nm15处理。培养一定时间后,分析它们对细胞的抑制生长作用。本发明发现相比于GFP-WT,GFP-Nm15具有更显著的抑制细胞生长作用,该作用虽然没有KPT-330强,但是GFP-Nm15的核输出抑制作用却是非常的显著。同时将CRM1的核输出作用与细胞的生长抑制作用一起分析,分析结果图如图13所示,图13中X轴表示细胞存活率,Y轴表示CRM1介导的NES出核率,数据均来自上图HeLa和A549细胞实验分析所得。由图13可知细胞的存活越高,其CRM1的出核运输效率越低,即细胞存活率与NES的出核率呈现负相关作用。也就是说,当抑制剂对CRM1核输出抑制作用越强的时候,其对细胞的生长抑制作用越明显。
实施例7 Nm15的进一步优化,获得最优Nm42
为了得到亲和力更高的NES,基于Nm15的CRM1的结构又优化设计了Nm42:DGYIEELIRMFGKLSIHDD(SEQ ID NO:10),通过ITC和PD的亲和力验证Nm42与CRM1亲和力,结果见图14。图14左图中GST-Nm15与GST-Nm42均在MBP-MVM的竞争下分别PD相同量CRM1,之后通过SDS/PAGE和考马斯亮蓝分析PD下的CRM1条带亮度。图14右图为ITC滴定实验分析Nm42与CRM1的亲和力,其Kd值约22nM。由图14可知,在与CRM1的亲和力上,Nm42比Nm15稍高。
同时在结构上也观察到,Nm42与CRM1的相互作用更偏疏水性,结果见图15。
实施例8 iDPP/DNA纳米复合物的获得及活性实验
考虑到现优化的NES肽在细胞穿模能力以及进入细胞后的稳定性上较差,通过常规分子克隆方法构建pEGFP-Nm15和pEGFP-Nm42质粒,将构建好的pEGFP-Nm15和pEGFP-Nm42质粒分别与iDPP纳米粒以1:25的摩尔比轻柔混合,室温30分钟孵育,得到iDPP/DNA纳米复合物,如图16所示。图16中mPEG-PLA为嵌段共聚物单甲氧基聚乙二醇-聚左旋丙交酯,C18-PEG-iRGD为肿瘤靶向肽,DOTAP为阳性2,3-二油氧基丙基)三甲基氯化铵(MSDS)。
用动态光散色检测iDPP/DNA纳米颗粒复合物直径约193nm,电位约2.12mV。
iDPP/DNA纳米复合物的细胞存活实验:将B16-F10细胞接种在96孔板中(5000个细胞/孔),并孵育24h,加入不同的iDPP/pEGFP-NES纳米复合物和转染试剂Turbo-2000,,在无血清培养基中孵育6个小时,之后换成完全培养基再在37℃条件下培养48个小时。然后在每个孔里加入20μL的MTT,并在黑暗环境下处理4个小时。最后直接在570nm波长下检测各孔的吸收值,以计算细胞存活率。通过MTT实验检测到对照组细胞存活率为96%,经iDPP/pEGFP-Nm15处理后的细胞存活率为66%,经iDPP/pEGFP-Nm42处理后的细胞存活率为61%,由此可知,经iDPP/pEGFP-Nm42处理后的细胞,其存活能力最低,结果见图17。也就是说在体内,iDPP/pEGFP-Nm42纳米复合物可有效抑制B16-F10细胞的生长。
实施例9 iDPP/Nm42纳米复合物的抗黑色素瘤活性分析和安全评估
(1)iDPP/DNA纳米复合物具有很高的肿瘤靶向效应-小鼠活体成像法
先对雌性C57BL/6小鼠进行静脉注射5×10
(2)iDPP/Nm42在体内的抗肿瘤活性分析
用以上方法建立的黑色素瘤小鼠模型,培育6天后,将它们平均分成4组。然后分别用iDPP/WT、iDPP/Nm42纳米复合物进行静脉注射处理,每两天注射一次,每次注射5ug,共注射4次。并每天记录小鼠的肿瘤体积和体重。15天后,将所有小鼠进行颈椎错位处死,并将小鼠肿瘤组织进行拍照,称重,同时将小鼠重要的器官(心,肝,肺,脾,肾等)保存到4%的多聚甲醛里。之后通过分析不同处理后,小鼠实体瘤的重量和体积的变化,来分析iDPP/Nm42在小鼠体内的抗肿瘤活性,结果见图19。图19左图表示相比对照组,经iDPP/Nm42纳米复合物处理后,小鼠肿瘤在重量上明显减轻。图19右图表示,经iDPP/Nm42纳米复合物处理后,小鼠肿瘤在体积上增长速率明显得到了抑制。
(3)iDPP/Nm42的安全评估
对小鼠进行以上不同处理后,对其血清的丙氨酸转氨酶(ALT)、天冬氨酸转氨酶(AST)和肌酐(CREA)的水平进行生化分析。同时也通过H&E染色对小鼠的重要器官进行了分析,最终判断iDPP/Nm42纳米复合物的安全性,结果见图20。由图20可知,iDPP/Nm42没有对小鼠的重要器官产生生理病变。同时观察小鼠的体重也没有大的波动(结果见图21),ALT、AST和CREA的水平也没有受影响(结果见图22,22-1、22-2和22-3分别为iDPP/Nm42对小鼠ALT、AST和CREA水平的影响结果图)。也就是说iDPP/Nm42纳米复合物在有效抗肿瘤的同时也对小鼠本身无毒性。
实施例10 CRM1的C528S突变细胞不会对多肽抑制剂产生耐药性
将不同浓度梯度(1nM-1uM)的KPT-330或Nm42加入到HeLa稳转细胞系中(CRM1 WT或C528S),每个浓度做至少6个重复。然后处理细胞,分析细胞存活能力,结果见图23。图23-1为KPT-330对CRM1的C528S突变细胞的耐药性分析结果,图23-2为Nm42对CRM1的C528S突变细胞的耐药性分析结果。由图23可知,KPT-330对CRM1-C528S的HeLa细胞的药效减弱,而经肽Nm42处理后,其抗肿瘤能力并没有因发生了C528S的突变而产生耐药性。
上述研究多肽对动物活性影响的实验中,本发明使用了基因治疗,但多肽的使用不限于基因疗法,也可以进一步优化多肽后,直接注射,或采用纳米包装多肽后注射,或者与其他制剂/药物联合使用。
序列表
<110> 四川大学
<120> 一种非共价靶向CRM1的多肽及其应用
<160> 10
<170> SIPOSequenceListing 1.0
<210> 1
<211> 19
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 1
Gly Ser Thr Val Glu Asp Met Thr Lys Lys Phe Gly Thr Leu Thr Ile
1 5 1015
His Asp Thr
<210> 2
<211> 19
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 2
Gly Ser Thr Val Asp Glu Leu Thr Lys Lys Phe Gly Thr Leu Thr Ile
1 5 1015
His Asp Thr
<210> 3
<211> 19
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 3
Gly Ser Thr Val Glu Asp Leu Thr Lys Lys Phe Gly Thr Leu Thr Ile
1 5 1015
His Asp Thr
<210> 4
<211> 20
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 4
Gly Ser Thr Val Asp Glu Met Thr Lys Lys Phe Gly Thr Leu Thr Ile
1 5 1015
His Asp Asp Asp
20
<210> 5
<211> 21
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 5
Gly Ser Asp Asp Thr Val Asp Glu Met Thr Lys Lys Phe Gly Thr Leu
1 5 1015
Thr Ile His Asp Thr
20
<210> 6
<211> 22
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 6
Gly Ser Asp Asp Thr Val Asp Glu Leu Thr Lys Lys Phe Gly Thr Leu
1 5 1015
Thr Ile His Asp Asp Asp
20
<210> 7
<211> 22
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 7
Gly Ser Asp Asp Thr Val Glu Asp Met Thr Lys Lys Phe Gly Thr Leu
1 5 1015
Thr Ile His Asp Asp Asp
20
<210> 8
<211> 22
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 8
Gly Ser Asp Asp Thr Val Glu Asp Leu Thr Lys Lys Phe Gly Thr Leu
1 5 1015
Thr Ile His Asp Asp Asp
20
<210> 9
<211> 19
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 9
Gly Ser Thr Val Asp Glu Met Thr Lys Lys Phe Gly Thr Leu Thr Ile
1 5 1015
His Asp Thr
<210> 10
<211> 19
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 10
Asp Gly Tyr Ile Glu Glu Leu Ile Arg Met Phe Gly Lys Leu Ser Ile
1 5 1015
His Asp Asp
一种非共价靶向CRM1的多肽及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0