

IPC分类号 : C12N9/00I,C12N15/52I,C12N15/70I,C12P13/04I,C12N1/21I,C12R1/19N









专利摘要
本发明公开一种侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用。本发明从侧耳类食用菌平菇、白灵菇、杏鲍菇出发,克隆并功能鉴定了这三种侧耳类食用菌麦角硫因合成编码相关基因,其核苷酸序列分别如SEQIDNO:2、4、6、8、10、12所示,分别与pET‑32a(+)表达载体连接并在大肠杆菌中实现了异源表达,并实现了体外合成麦角硫因。本发明首次阐明了侧耳类食用菌麦角硫因生物合成途径,表明其具有更简洁的合成通路,即由Pegt1、Pegt2两个基因参与麦角硫因的合成,同时,本发明构建的基因工程细胞安全稳定,生产周期短,显示出其在麦角硫因的应用开发中的重大价值。
权利要求
1.侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用,其特征在于:
所述的侧耳类食用菌为平菇、白灵菇或杏鲍菇;
所述的平菇麦角硫因合成酶PEGT1的氨基酸序列如SEQ ID NO:1所示;
所述的平菇麦角硫因合成酶PEGT2的氨基酸序列如SEQ ID NO:3所示;
所述的白灵菇麦角硫因合成酶PEGT1的氨基酸序列如SEQ ID NO:5所示;
所述的白灵菇麦角硫因合成酶PEGT2的氨基酸序列如SEQ ID NO:7所示;
所述的杏鲍菇麦角硫因合成酶PEGT1的氨基酸序列如SEQ ID NO:9所示;
所述的杏鲍菇麦角硫因合成酶PEGT2的氨基酸序列如SEQ ID NO:11所示。
2.根据权利要求1所述的应用,其特征在于:
所述的侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在体外联合生物合成麦角硫因中的应用。
3.根据权利要求1或2所述的应用,其特征在于:
所述的平菇麦角硫因合成酶PEGT1编码基因的核苷酸序列如SEQ ID NO:2所示;
所述的平菇麦角硫因合成酶PEGT2编码基因的核苷酸序列如SEQ ID NO:4所示;
所述的白灵菇麦角硫因合成酶PEGT1编码基因的核苷酸序列如SEQ ID NO:6所示;
所述的白灵菇麦角硫因合成酶PEGT2编码基因的核苷酸序列如SEQ ID NO:8所示;
所述的杏鲍菇麦角硫因合成酶PEGT1编码基因的核苷酸序列如SEQ ID NO:10所示;
所述的杏鲍菇麦角硫因合成酶PEGT2编码基因的核苷酸序列如SEQ ID NO:12所示。
4.侧耳类食用菌麦角硫因合成酶PEGT1,其特征在于:
所述的侧耳类食用菌为平菇、白灵菇或杏鲍菇;
所述的平菇麦角硫因合成酶PEGT1的氨基酸序列为如SEQ ID NO:1所示氨基酸序列构成的蛋白;
所述的白灵菇麦角硫因合成酶PEGT1的氨基酸序列为如SEQ ID NO:5所示氨基酸序列构成的蛋白;
所述的杏鲍菇麦角硫因合成酶PEGT1的氨基酸序列为如SEQ ID NO:9所示氨基酸序列构成的蛋白。
5.权利要求4所述的侧耳类食用菌麦角硫因合成酶PEGT1的编码基因,其特征在于:所述的平菇麦角硫因合成酶PEGT1的编码基因的核苷酸序列为如SEQ ID NO:2所示的核苷酸序列;
所述的白灵菇麦角硫因合成酶PEGT1的编码基因的核苷酸序列为如SEQ ID NO:6所示的核苷酸序列;
所述的杏鲍菇麦角硫因合成酶PEGT1的编码基因的核苷酸序列为如SEQ ID NO:10所示的核苷酸序列。
6.侧耳类食用菌麦角硫因合成酶PEGT2,其特征在于:其氨基酸序列为如下所示:SEQID NO:7所示氨基酸序列构成的蛋白;
所述的侧耳类食用菌为白灵菇。
7.权利要求6所述的侧耳类食用菌麦角硫因合成酶PEGT2的编码基因,其特征在于:其核苷酸序列为如下所示:SEQ ID NO:8所示的核苷酸序列。
8.侧耳类食用菌麦角硫因合成酶PEGT2的编码基因,其特征在于:
所述的侧耳类食用菌为平菇或杏鲍菇;
所述的平菇麦角硫因合成酶PEGT2的编码基因的核苷酸序列为SEQ ID NO:4所示的核苷酸序列;
所述的杏鲍菇麦角硫因合成酶PEGT2的编码基因的核苷酸序列为SEQ ID NO:12所示的核苷酸序列。
9.含有权利要求5或7或8所述的编码基因的重组载体、表达盒或重组菌。
10.权利要求9所述的重组载体、表达盒或重组菌在合成麦角硫因中的应用。
11.根据权利要求10所述的应用,其特征在于:
构建含有权利要求5或7或8编码基因的重组表达载体,将重组表达载体转化至大肠杆菌BL21(DE3)中,将获得的基因工程细菌进行诱导表达,用PBS洗涤收集诱导表达后的菌体,最后将含有PEGT1工程菌与PEGT2工程菌混合,用PBS溶解混匀后添加反应底物破碎后反应,得到含有麦角硫因的发酵液。
说明书
技术领域
本发明属于合成生物学领域,具体涉及侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用。
背景技术
麦角硫因(Ergothioneine,简称EGT)是一种稀有的天然手性氨基酸类强抗氧化剂,主要在细菌和真菌等微生物体内合成,其具有独特的生物学功能和药理作用,在食品和医药等行业极具应用前景。
但背景技术方面存在以下几点不足:
(1)生物合成产量普遍低下且合成过程繁琐,已报道的来自耻垢分枝杆菌的参与麦角硫因合成的五基因EgtA、B、C、D、E异源表达后虽然合成麦角硫因产量达到24mg/L,但耻垢分枝杆菌具有一定的生物危害性,且麦角硫因生物合成过程周期长。
(2)以生物为原料直接提取成本高,产量低,且来源有限。
(3)由于麦角硫因结构特性,化学合成难度大,且合成原料昂贵。
(4)直接利用食用菌菌丝体深层发酵获得麦角硫因是近年来生产麦角硫因的主要手段,但生产周期长及其生产成本高和产率低等问题严重限制麦角硫因的大规模生产。
(5)已有文献报道表明,真菌中首次EGT生物合成通路2014年在粗糙脉孢菌EGT生物合成途径中完成解析,这为更多真菌EGT生物合成相关酶基因的发现和EGT生物合成通路的解析提供了更好的研究基础。真菌中具有更简洁的麦角硫因生物合成通路,食用菌作为很好的药食同源真菌,是麦角硫因合成酶基因的良好来源,目前,侧耳类食用菌麦角硫因合成酶编码基因尚未有文献报道。
发明内容
为了克服现有技术的缺点与不足,本发明的目的在于提供一种侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用。
本发明的另一目的在于提供侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2。
本发明的另一目的在于提供上述侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因Pegt1、Pegt2。
本发明的目的通过下述技术方案实现:
本发明提供一种侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用。
优选的,侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在体外联合生物合成麦角硫因中的应用。
优选的,所述的侧耳类食用菌为平菇、白灵菇或杏鲍菇。
所述的平菇麦角硫因合成酶PEGT1(PBEGT1)的氨基酸序列如SEQ ID NO:1所示,其编码基因(PBegt1)的核苷酸序列如SEQ ID NO:2所示;
所述的平菇麦角硫因合成酶PEGT2(PBEGT2)的氨基酸序列如SEQ ID NO:3所示,其编码基因(PBegt2)的核苷酸序列如SEQ ID NO:4所示;
所述的白灵菇麦角硫因合成酶PEGT1(PtEGT1)的氨基酸序列如SEQ ID NO:5所示,其编码基因(Ptegt1)的核苷酸序列如SEQ ID NO:6所示;
所述的白灵菇麦角硫因合成酶PEGT2(PtEGT2)的氨基酸序列如SEQ ID NO:7所示,其编码基因(Ptegt2)的核苷酸序列如SEQ ID NO:8所示;
所述的杏鲍菇麦角硫因合成酶PEGT1(PeEGT1)的氨基酸序列如SEQ ID NO:9所示,其编码基因(Peegt1)的核苷酸序列如SEQ ID NO:10所示;
所述的杏鲍菇麦角硫因合成酶PEGT2(PeEGT2)的氨基酸序列如SEQ ID NO:11所示,其编码基因(Peegt2)的核苷酸序列如SEQ ID NO:12所示;
SEQ ID NO:1、3、5、7、9、11所示的氨基酸序列经取代和/或缺失和/或添加一个或几个氨基酸残基且功能相同的衍生蛋白也属于本发明的保护范围。
SEQ ID NO:2、4、6、8、10、12所示的核苷酸序列经取代和/或缺失和/或添加一个或几个核苷酸且表达相同功能蛋白的核苷酸序列也属于本发明的保护范围。
本发明还提供侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2,具体内容为(1)或(2):
(1)SEQ ID NO:1、3、5、7、9、11所示氨基酸序列构成的蛋白;
(2)SEQ ID NO:1、3、5、7、9、11所示的氨基酸序列经取代和/或缺失和/或添加一个或几个氨基酸残基且功能相同的衍生蛋白。
本发明还提供侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因,其为(1)或(2):
(1)SEQ ID NO:2、4、6、8、10、12所示的核苷酸序列;
(2)SEQ ID NO:2、4、6、8、10、12所示的核苷酸序列经取代和/或缺失和/或添加一个或几个核苷酸且表达相同功能蛋白的核苷酸序列;
含有所述编码基因的重组载体、表达盒、重组菌也属于本发明的保护范围。
所述的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2或其编码基因在制备含有侧耳类食用菌麦角硫因合成酶编码基因的重组载体、表达盒、重组菌中的应用。
所述的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2或其编码基因在制备含有麦角硫因的发酵液的应用,所述的应用是采用:构建含有编码所述基因的重组表达载体,将重组表达载体转化至大肠杆菌BL21(DE3)中,将获得的基因工程细菌进行诱导表达,用PBS洗涤收集诱导表达后的菌体,最后将含有PEGT1工程菌与PEGT2工程菌混合,用PBS溶解混匀后添加反应底物破碎后反应2小时,得到含有麦角硫因的发酵液。
本发明还提供获取侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA序列的特异性引物对,所用引物均由广州市天一辉远基因科技有限公司合成。
本发明提供的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因是从侧耳类食用菌平菇、白灵菇、杏鲍菇中克隆获得的麦角硫因合成酶基因,其能够在S-腺苷蛋氨酸(SAM)、Fe
侧耳类食用菌麦角硫因合成酶的发现丰富了麦角硫因合成酶的多样性,此外能够在一定程度上解决该化合物的来源及资源问题。
本发明相对于现有技术具有如下的优点及效果:
本发明从侧耳类食用菌平菇、白灵菇、杏鲍菇出发,克隆并功能鉴定了这三种侧耳类食用菌麦角硫因合成编码相关基因,其核苷酸序列分别如SEQ ID NO:2、4、6、8、10、12所示,分别与pET-32a(+)表达载体连接并在大肠杆菌中实现了异源表达,并实现了体外合成麦角硫因。麦角硫因具有重要的应用价值,本发明首次阐明了侧耳类食用菌麦角硫因生物合成途径,表明其具有更简洁的合成通路,即由Pegt1、Pegt2两个基因参与麦角硫因的合成,同时,本发明构建的基因工程细胞安全稳定,生产周期短,显示出其在麦角硫因的应用开发中的重大价值。
附图说明
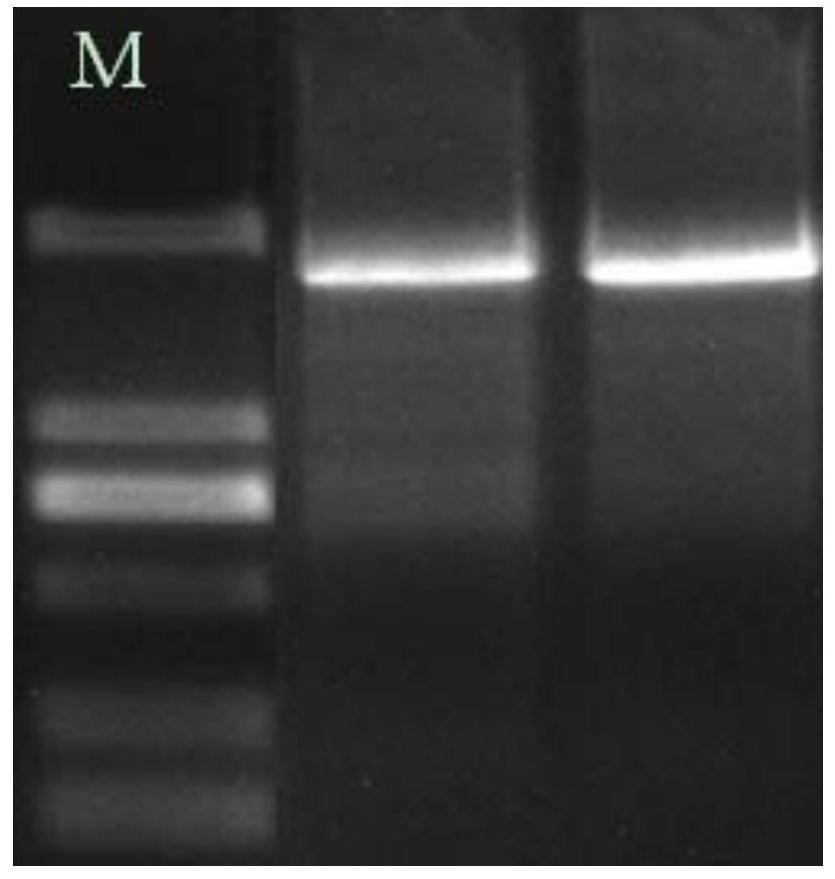
图1为侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因的PCR琼脂糖凝胶电泳图,其中,a为侧耳类食用菌提取的RNA电泳图,泳道1、2:平菇,泳道3、4:白灵菇,泳道5、6:杏鲍菇;b为RT-PCR扩增得到的侧耳类食用菌麦角硫因合成酶编码基因cDNA电泳图,泳道M:DNAMarker3,泳道1:PBegt1,泳道2:PBegt2,泳道3:Ptegt1,泳道4:Ptegt2,泳道5:Peegt1,泳道6:Peegt2。
图2为表达平菇麦角硫因合成酶PBEGT1、PBEGT2编码基因的大肠杆菌表达质粒pET-32a-PBegt1、pET-32a-PBegt2的结构示意图。
图3为表达白灵菇麦角硫因合成酶PtEGT1、PtEGT2编码基因的大肠杆菌表达质粒pET-32a-Ptegt1、pET-32a-Ptegt2的结构示意图。
图4为表达杏鲍菇麦角硫因合成酶PeEGT1、PeEGT2编码基因的大肠杆菌表达质粒pET-32a-Peegt1、pET-32a-Peegt2的结构示意图。
图5为侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因表达载体的诱导表达蛋白SDS-PAGE电泳图,其中,泳道1:PBEGT1,泳道2:PBEGT2,泳道3:PtEGT1,泳道4:PtEGT2,泳道5:PeEGT1,泳道6:PeEGT2。
图6为杏鲍菇麦角硫因合成酶PeEGT编码基因诱导表达后生物合成麦角硫因反应产物HPLC鉴定图。
图7为平菇、白灵菇麦角硫因合成酶PBEGT和PtEGT编码基因诱导表达后生物合成麦角硫因反应产物HPLC鉴定图。
具体实施方式
下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。
下列实施例中未注明具体实验条件的试验方法,通常按照常规实验条件或按照制造厂所建议的实验条件。所使用的材料、试剂等,如无特殊说明,为从商业途径得到的试剂和材料。
实施例1
侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA序列的获取及生物信息学分析:
利用已报道的粗糙脉孢菌麦角硫因合成酶Nc-EGT1(Protein ID:XP_956324.3)、Nc-EGT2(Protein ID:XP_001728131.1)氨基酸序列(Bioinformatic and biochemicalcharacterizations of C-S bond formation and cleavage enzymes in the fungusNeurospora crassa ergothioneine biosynthetic pathway[J].Organic Letters,2014,16(20):5382-5385.)放在NCBI中运用BLASTP与平菇进行同源性搜索,找到平菇中与其相似的对应两个蛋白,从而找到平菇中对应蛋白的cDNA序列,利用平菇的两个cDNA序列运用BLASTN分别跟侧耳类食用菌白灵菇和杏鲍菇基因组进行分析比对,进而找到相应的侧耳类食用菌白灵菇和杏鲍菇麦角硫因合成酶PEGT1、PEGT2编码基因。利用PDB培养基(土豆去皮200g/L,葡萄糖20g/L,MgSO4·7H2O g/L,KH2PO4g/L,自然pH)摇菌收集侧耳类食用菌菌丝体进行RNA提取(见图1a),以提取的质量好的侧耳类食用菌RNA为模板,采用全式金公司的TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix反转录试剂盒的方法合成cDNA链,用分析得到的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因为引物设计模板,利用primer premier5设计引物分别扩增侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA序列(如SEQ ID NO:2、4、6、8、10、12),具体扩增引物如下表1(测序结果发现扩增的个别序列与原先基因组为模板设计的引物有个别碱基不匹配,已根据实际获取的测序序列对引物进行优化)。
表1.侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA扩增引物
对扩增得到的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA利用Swiss-Model进行氨基酸序列比对、蛋白三维结构的同源模拟和结构合理性在线分析发现,侧耳类食用菌麦角硫因合成酶PEGT1同时具有类似于亚砜合酶EgtB活性和组氨酸特异性甲基转移酶EgtD活性。而侧耳类食用菌麦角硫因合成酶PEGT2则具有类属于Nc-EGT2的半胱氨酸亚砜裂解酶活性,根据分析结果,可初步推断侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2能够合成麦角硫因。
实施例2
侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA序列表达载体构建:
(1)菌株和质粒
(2)利用PDB培养基摇菌收集侧耳类食用菌菌丝体进行RNA提取,RNA提取具体操作过程如下:
1)将新鲜收集的真菌菌丝置于无RNA酶的液氮预冷的研钵中,加入适量的液氮,用无RNA酶的研磨杵迅速将菌丝研磨成粉末,迅速取约80mg的粉末于无RNA酶的1.5mL离心管中。
2)立即加入300μL抽提缓冲液(分别吸取200μL 10xSTE Buffer(称取3.025gTris,0.146g EDTA,2.9g NaCl于50mL烧杯中,先加约30mL ddH2O溶解,再用盐酸调pH至8.0,定容至50mL后高压蒸汽灭菌)、100μL 10%SDS(5gSDS溶解于50mL ddH2O)和700μL无菌去离子水于2mL离心管中,混合均匀)和600μL PCI(PCI由酚:氯仿:异戊醇以25:24:1的比例混合而成),将离心管置于振荡器中振荡30s。
3)在4℃、14000rpm条件下离心5min,用移液枪吸取上清于另一离心管中,加入等体积的PCI,在振荡器中振荡20s。
4)在4℃、14000rpm条件下离心5min,用移液枪小心吸取上清于另一离心管中,加入1/10体积的3M NaAc(pH5.2)(24.6g无水NaAc于烧杯中,加入约40mL去离子水搅拌溶解,加冰醋酸调pH至5.2,用去离子水定容至100mL),摇匀。
5)加入2-2.5倍体积的无水乙醇,置于振荡器中振荡混匀,于-20℃沉淀约40min。
6)在4℃、14000rpm条件下离心15min,弃去上清液。加入一定量的70%的乙醇洗涤沉淀,在4℃、14000rpm条件下离心5min,倒掉乙醇。
7)用移液枪尽量移去液滴,以去除洗涤乙醇,在真空干燥器中干燥5min或在超净工作台中干燥5min。
8)用10μL去RNase的ddH2O溶解沉淀,即得到提取的总RNA溶液,存于-80℃备用。
(3)将提取的质量好的侧耳类食用菌RNA为模板,采用全式金公司的TransScriptOne-Step gDNA Removal and cDNA Synthesis SuperMix反转录试剂盒的方法合成cDNA链,具体操作详见说明书。
(4)以合成的cDNA链为模板,利用设计的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA扩增引物(见表1),采用高保真酶KOD-Fx进行PCR扩增,扩增体系为25μL,反应程序为:2xPCR Buffer 12.5μL,2mM dNTPs 5μL,Primer F 0.75μL,Primer R 0.75μL,Template 0.5μL,KOD-Fx 0.5μL,ddH2O补足至25μL。PCR程序为:94℃预变性2min;98℃变性10s,Pegt1引物退火57℃,30s,68℃延伸2min34s,30个循环,Pegt2引物退火60℃,30s,68℃延伸1min14s,30个循环,程序结束以后用1%琼脂糖凝胶电泳检测条带大小,结果见图1b,其中M代表DNA Marker3,1、2、3、4、5、6分别代表PBegt1、PBegt2、Ptegt1、Ptegt2、Peegt1、Peegt2电泳条带,从图中可以看出,扩增得到的条带与预期的条带大小相一致(侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA分别约2562bp和1245bp),由此初步判定侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA扩增成功。
(5)侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因pCloneEZ-TOPO克隆载体的构建
将PCR扩增好的目的条带分别与pCloneEZ-TOPO载体连接,具体操作详见说明书,将连接产物转化大肠杆菌DH5a,涂布在添加氨苄青霉素(Amp:100μg/mL)的LB固体平板进行筛选,37℃恒温培养箱过夜培养至长出单菌落,挑取单克隆进行菌落PCR验证,选出阳性克隆进行DNA测序验证。
(6)摇菌提取成功构建的侧耳类食用菌PEGT1、PEGT2编码基因pCloneEZ-TOPO克隆载体质粒及pET-32a(+)质粒,选择EcoRI作为酶切位点,采用EcoRI限制性内切酶对pET-32a(+)进行单酶切线性化(反应体系:pET-32a(+)<5μg,10xFastDigest Buffer 5μL,FastDigestEnzyme 3μL,补充ddH2O至50μL),37℃反应25分钟后85℃反应5分钟使酶失活并进行产物回收纯化。采用CE Design V1.04软件设计带有pET-32a(+)线性化同源臂的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA扩增引物,具体引物序列如下表2。以侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因pCloneEZ-TOPO克隆载体质粒为模板,采用高保真酶PrimerSTAR Max Premix(2x)进行PCR增,PCR体系为25μL,反应程序为:PrimerSTAR Max Premix(2x)12.5μL,Primer F 0.75μL,Primer R 0.75μL,Template 0.2μL,ddH2O补足至25μL。PCR程序为:98℃变性10s,加同源臂Pegt1引物退火60℃,30s,68℃延伸2min35s,30个循环,加同源臂Pegt2引物退火60℃,30s,68℃延伸1min15s,30个循环,程序结束以后用1%琼脂糖凝胶电泳检测条带大小,并进行产物回收纯化。采用ClonExpressII One Step Cloning Kit同源重组试剂盒将带同源臂的侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA连接到EcoRI单酶切线性化的pET-32a(+)载体上(结构示意图见图2、3和4),具体操作详见说明书;将连接产物转化大肠杆菌BL21(DE3),涂布在添加氨苄青霉素(Amp:100μg/mL)的LB(胰化蛋白胨10g/L,酵母提取为5g/L,NaCl 10g/L,pH7.4,琼脂粉20g/L(固体))固体平板进行筛选,37℃恒温培养箱过夜培养至长出单菌落,挑取单克隆进行PCR验证,选出阳性克隆进行DNA测序验证。
表2.带同源臂侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2编码基因cDNA扩增引物
实施例3
侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2的蛋白诱导表达分析:
按照1%(v/v)的接种量将构建成功的工程菌液接种到50mL LB液体培养基中,37℃,220rpm培养至OD600nm=0.6~0.8左右,加入终浓度为0.6mM的IPTG,在20℃条件下诱导表达16h。利用pH7.4的PBS(称取8gNaCl、0.2gKCl、1.42gNa2HPO4、0.27gKH2PO4于烧杯中,加入约800mL的去离子水充分溶解,用盐酸调pH至7.4后用ddH2O定容至1L高压蒸汽灭菌)洗涤诱导表达后的工程菌株后,取适量的菌体溶于PBS后与2x蛋白上样Buffer(添加0.5mL 1MpH6.8的Tris-HCl(称取121.1g Tris置于1L烧杯中,加入约800mL的去离子水,充分搅拌溶解,用浓盐酸调节pH至6.8后定容至1L,高压蒸汽灭菌,室温保存)、0.2g SDS(十二烷基硫酸钠)、10mg BPB(溴酚蓝)、1mL甘油于10mL离心管中,加去离子水定容至5mL,500μL小份分装于离心管后,使用前每小份添加10μLβ-巯基乙醇)按1:1比例混匀,沸水浴10分钟后冷却至室温,利用7.5%SDS-PAGE(分离胶的制备:取分离胶贮存液1.25mL,Tris-HCl缓冲液(pH8.8)0.625mL,10%SDS 0.05mL,10%过硫酸铵0.025mL,1%TEMED 0.5mL,去离子水2.55mL。充分混匀后,用1mL枪头加至两块玻璃板之间,高度为距样品模梳齿下缘约1cm,并赶尽胶表面气泡,继续加入800μL蒸馏水使胶面平整,并隔绝空气。45分钟后,待胶块凝固后倒掉蒸馏水,并用吸水纸吸去多余的水。浓缩胶的制备:取浓缩胶贮存液0.6mL,Tris-HCl缓冲液(pH6.8)0.25mL,10%SDS 0.02mL,10%过硫酸铵0.01mL,1%TEMED 0.4mL,去离子水0.92mL。充分混匀后,用1mL枪头加至两块玻璃板之间分离胶上层,轻轻插入样品模梳齿,待胶凝固后,拔出梳齿)检测重组蛋白是否正确表达。结果如图5所示,其中M代表PageRuler PrestainedProtein Ladder,1、2、3、4、5、6分别代表PBEGT1、PBEGT2、PtEGT1、PtEGT2、PeEGT1、PeEGT2重组蛋白SDS-PAGE电泳条带,从图中可以看出,表达的重组蛋白分别与预期条带大小一致(侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2重组蛋白分别约114.7kDa和64.1kDa),说明构建的表达载体能够有效以重组蛋白形式分别表达侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2。
实施例4
侧耳类食用菌麦角硫因合成酶PEGT1、PEGT2体外酶活测试分析:
1)按照1%(v/v)的接种量将构建成功的工程菌液和对照菌株(不含目的基因,含质粒pET-32a(+))接种到50mL LB液体培养基中,37℃,220rpm培养至OD600nm=0.6~0.8左右,加入终浓度为0.6mM的IPTG,在20℃条件下诱导表达16h。
2)加入30μL IPTG,将IPTG的终浓度控制在0.6mM,在20℃诱导表达16h;
3)利用pH7.4的PBS分别洗涤收集细胞,最后将来源于同一菌种的PEGT1和PEGT2工程菌株混合并溶解在10mL PBS中,加入反应底物以控制最终浓度为:20mM His,31mg;20mMCys,24.3mg;5mM SAM,20mg;10mM Fe
4)混匀,超声破碎细菌,破碎条件:功率33%,约250W;总超声时间6min,工作3s,间隔3s,报警温度40℃;
5)超声结束后,将反应体系放置在36℃下反应2小时,期间每隔20分钟混匀一次;
6)反应产物利用0.22μm水系滤膜过滤后用于HPLC检测,其中平菇和白灵菇麦角硫因合成酶PBEGT和PtEGT体外酶活测试采用液相检测条件为:检测波长257nm,流速1mL/min,流动相(乙腈:20mM,pH6.0乙酸铵=85:15),单次样品分析时间60min,杏鲍菇麦角硫因合成酶PeEGT体外酶活测试采用液相检测条件为:检测波长257nm,流速1mL/min,流动相(乙腈:水=80:20),单次样品分析时间60min。
结果如图6和7所示,图6中,PeEGT1和PeEGT2酶促反应产物在41.700min处出峰,与EGT标准品出峰时间41.614min基本相一致。图7中,PBEGT1和PBEGT2酶促反应产物以及PtEGT1和PtEGT2酶促反应产物出峰时间分别为46.482min和46.459min,也基本与EGT标准品出峰时间46.863相一致。图6和图7中,空白对照组在EGT标品出峰时间处均没有检测到产物出现,由此可以看出,克隆表达的侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2能够在体外联合生物合成麦角硫因。
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
序列表
<110> 华南农业大学
<120> 侧耳类食用菌麦角硫因合成酶PEGT1和PEGT2在合成麦角硫因中的应用
<160> 36
<170> SIPOSequenceListing 1.0
<211> 853
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 1
Met Ala Ile Gln Ile Val Asp Val Gln Thr Leu Asn Gln Ser Arg His
1 5 1015
Leu Asp Leu Gln Ser Asp Leu Ser Gln Gln Leu Val Asp Gly Leu Thr
202530
Arg Pro His Gly Glu Lys Glu Ile Pro Thr Val Leu Leu Tyr Asp Glu
354045
Arg Gly Leu Arg Leu Tyr Asp Ala Ile Thr Thr Glu Val Ser Glu Tyr
505560
Tyr Leu Phe Gly Ala Glu Glu Glu Ile Leu Lys Asn Lys Ala Asp Glu
65707580
Ile Val Arg Thr Met His Ser Gly Leu Gly Asp Gly Asp Pro Asp Ser
859095
Glu Val Val Leu Glu Leu Gly Ala Gly Ser Leu Arg Lys Thr Ser His
100 105 110
Ile Leu Ala Gly Leu Ser Arg Ile Val Pro Ser Gln Cys Asp Thr Ala
115 120 125
Pro Ile Thr Tyr Tyr Ala Leu Asp Leu Glu Glu Arg Glu Leu Gln Arg
130 135 140
Val Leu Asp Ser Ile Ser Asn Ser Ser Val Gly Glu Met Leu Arg Gly
145 150 155 160
Lys Val Glu Thr Lys Gly Leu Trp Gly Thr Tyr Glu Asp Gly Leu Lys
165 170 175
Phe Val Glu Asp Glu Gly Ser Pro Ile Ser Ser His Ser Asn Ser Pro
180 185 190
Thr Ser Gln Ser Ser Arg Arg Glu Leu Gly Pro Pro Ser Pro Thr Pro
195 200 205
Arg Ser Gly Ser Ser Ser Pro Leu His Ile Leu Phe Leu Gly Ser Ser
210 215 220
Leu Gly Asn Phe Asp Arg Lys Asp Ser Val Lys Phe Leu Gln Ser Met
225 230 235 240
Pro Leu Arg Pro Gly Ser Gly Asp Thr Leu Leu Ile Gly Leu Asp His
245 250 255
Asp Asn Asp Lys Thr Leu Ile Glu Glu Ala Tyr Asn Asp Arg Lys Gly
260 265 270
Tyr Thr Lys Thr Phe Ile Met Asn Gly Leu Arg Ala Ala Gly Arg Ala
275 280 285
Leu Gly Asn Glu Asp Met Phe Glu Glu Asp Lys Trp Glu Tyr Val Asn
290 295 300
Arg Tyr Asn Glu Ala Glu Arg Arg His Glu Ala Tyr Tyr Lys Ser Lys
305 310 315 320
Cys Pro Gln Lys Leu Glu Asp Pro Lys Ala Glu His Gly Tyr Glu Phe
325 330 335
Leu Glu Asp Glu Leu Val Lys Ile Glu Val Ser Tyr Lys Tyr Ser Glu
340 345 350
Thr Asp Ala Tyr Ser Leu Phe Thr Asp Ser Gly Leu Arg Pro Ile Gln
355 360 365
Arg Trp Thr Asp Ser Ala Thr Gln Tyr Ser Leu Trp Leu Leu Glu Arg
370 375 380
Pro Pro Phe Met Phe Pro Leu Leu Lys Ser Pro Ile Ala Phe Asn Gly
385 390 395 400
Leu Gly Glu Ile Val Lys Asn Phe Pro Leu Ser Asn Thr Pro Phe Gly
405 410 415
Val Pro Ser Pro Gln Glu Trp Ser Asn Leu Trp Ala Ala Trp Asp Phe
420 425 430
Val Thr Leu Lys Met Ile Pro Pro Ser Met Leu His Val Lys Pro Ile
435 440 445
His Leu Arg His Ile Cys Leu Phe Tyr Leu Gly His Ile Pro Thr Phe
450 455 460
Leu Asp Ile His Leu Ser Arg Leu Leu Lys Glu Pro His Thr Glu Pro
465 470 475 480
Glu Ser Phe Lys Tyr Ile Phe Glu Arg Gly Ile Asp Pro Asn Val Asp
485 490 495
Asn Pro Ala Glu Cys His Pro His Ser Glu Val Pro Gln Asn Glu Gly
500 505 510
Asp Trp Pro Ser Leu Glu Ala Ile Leu Ser Phe Gln Ala Gln Val Arg
515 520 525
Ser Arg Leu Leu His Leu Tyr Asp Asp Val Leu Thr Gly Lys Lys Val
530 535 540
Leu Thr Arg Lys Val Cys Arg Ile Leu Ser Met Thr Phe Glu His Glu
545 550 555 560
Ala Phe His Ile Glu Thr Leu Leu Tyr Met Leu Leu Gln Ala Ala Gly
565 570 575
Asn Gly Thr Thr Pro Pro Pro Gly Phe Thr Pro Pro Pro Trp Ser Ser
580 585 590
Leu Ser Ala Asn Trp Asn Ala Leu Pro Ala Leu Glu Asn Thr Thr Val
595 600 605
Thr Leu Gly Pro Glu Ile Val Ala Leu Gly His Asp Asp Asp Glu Arg
610 615 620
Glu Asp Glu Asp Pro Ala Phe Ile Thr Asp Val Lys Asp His Glu Phe
625 630 635 640
Ala Trp Asp Asn Glu Asn Pro Lys Arg Gln Val Glu Val Lys Gln Phe
645 650 655
Lys Ile Glu Trp Arg Pro Val Thr Asn Gly Gln Phe Tyr Glu Phe Tyr
660 665 670
Lys Glu Tyr Lys Asp Asp Met Lys Leu Gln Leu Pro Ala Ser Trp Leu
675 680 685
Glu Thr Asp Asp Lys Met Met Val Arg Thr Leu Tyr Gly Pro Val Pro
690 695 700
Met Lys Val Ala Gln Asn Trp Pro Val Val Ile Ser Tyr Asp Gly Leu
705 710 715 720
Ser Ala Tyr Ala Ile Val Lys Gly Gly Arg Leu Pro Thr Glu Pro Glu
725 730 735
Leu Arg Leu Phe Leu Asp Lys Phe Glu Ser Gly Tyr Glu Gly Gly Ala
740 745 750
Asn Val Gly Phe Arg Asn Trp His Pro Val Pro Ala Thr Thr Gly Gly
755 760 765
Lys Lys Tyr Gly Gly Lys Gly His Asn Gly Gly Val Trp Glu Trp Thr
770 775 780
Ser Thr Thr Leu Thr Lys His Asp Gly Phe Glu Pro Ser Lys His Tyr
785 790 795 800
Pro Gly Phe Thr Ala Asp Phe Phe Asp Gly Cys His Asn Val Ile Leu
805 810 815
Gly Gly Ser Tyr Ala Phe Ile Pro Arg Met Ala Glu Arg Arg Ser Leu
820 825 830
Arg Asn Trp Tyr Gln Arg Asn Tyr Pro Tyr Ala Trp Thr Gly Gly Arg
835 840 845
Ile Val Tyr Asp Ile
850
<211> 2562
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 2
atggccattc aaatcgtcga cgtccagacg ctcaatcagt ccagacattt ggatctgcag 60
agcgacctct cccagcagct tgttgacggc ttaactcggc cgcatggcga aaaggaaatc 120
cccaccgtgc tcttatacga cgaacgaggg ttaagactct acgacgccat cacgaccgaa 180
gtctcggaat actacctctt tggcgccgag gaggaaatcc tcaagaataa agccgatgag 240
attgttcgga cgatgcacag tggattgggg gacggggatc cagactcgga ggtcgtcctc 300
gaattaggcg ccggttctct aaggaagacc tcccacattc tcgcagggct ttcacgcatc 360
gttcctagcc agtgcgatac agcgccgatc acctactacg ccctcgatct tgaagaacgc 420
gaacttcagc gagtactaga ctccatatca aactcaagcg ttggagagat gttgcgagga 480
aaagtagaaa cgaagggcct ctggggcact tacgaagacg ggctgaagtt tgtagaagac 540
gagggttctc ccatcagcag ccattcaaat tcccctacta gccaatcttc gagaagagag 600
ttggggccgc catcaccgac ccccagatcg ggatcgagtt ccccactgca tattctattc 660
ttaggttcgt cgttggggaa ctttgaccgc aaggatagcg tcaagttctt gcagtccatg 720
ccccttcgcc ctggctcagg cgatacgcta ctaattggtc tcgatcacga taacgataag 780
acgttgatcg aagaggcgta caatgaccgc aaaggatata cgaagacgtt catcatgaac 840
gggttgagag cggctggacg tgcgctagga aacgaggata tgtttgaaga agacaaatgg 900
gaatatgtta atagatataa tgaggcggag cgtcggcatg aggcatatta caagtccaag 960
tgtccccaga aactcgaaga tccgaaggcg gaacatggct acgagttcct tgaagatgaa 1020
ctagtcaaaa tcgaggtctc ctataagtac tcggaaaccg atgcctatag cctattcact 1080
gactcgggcc tccgccctat ccaacgttgg acagacagtg ccacccagta ctccctttgg 1140
ctactcgagc gtcccccctt tatgttcccc cttttgaagt ccccaatcgc cttcaatggc 1200
ctcggcgaga tcgtcaaaaa ctttcccctc tccaacacgc ccttcggcgt tccttcaccc 1260
caagaatggt caaacctgtg ggccgcttgg gacttcgtga ccttgaagat gattccaccc 1320
tcgatgctcc acgtgaagcc tatccacctg aggcacatat gcctatttta tctgggccat 1380
atcccgacgt tcttggacat ccatttgagt cgcttgctga aagaaccgca cacggagcct 1440
gaaagtttca agtatatctt tgagcgaggg attgatccta atgtggacaa cccggccgaa 1500
tgtcatcccc attcagaggt tccgcagaat gaaggcgatt ggccttcgct tgaagctata 1560
ttatcctttc aagcccaagt ccgctcaagg cttttgcatc tctacgacga tgtcttgaca 1620
ggaaagaagg tgttgacgcg aaaggtctgt cgcatattgt caatgacttt cgaacacgag 1680
gctttccaca ttgagacgtt gctctacatg cttttgcaag ccgccggaaa cgggactacc 1740
cctcccccag gattcacacc tccaccctgg agctcgcttt ccgcaaactg gaacgctctc 1800
ccagcgttgg aaaacacgac agttactctg ggtccggaaa tcgtagcatt aggccacgac 1860
gacgatgaaa gggaggatga agaccctgcg ttcatcactg acgtgaaaga tcacgaattt 1920
gcatgggata atgagaatcc caagcgtcag gtcgaagtca agcagttcaa gattgagtgg 1980
aggccggtca cgaatggcca gttctacgaa ttctacaaag agtataagga cgacatgaag 2040
ctccaattac cggccagttg gttggagaca gatgataaga tgatggttcg caccctgtac 2100
ggccctgtac ctatgaaagt ggcgcagaac tggccggtcg tcatttcgta cgatggcttg 2160
tctgcctatg cgatcgttaa aggcggaagg cttccaaccg agcctgagct tcgtctcttc 2220
ctagataaat ttgagagcgg ctacgaaggc ggcgcaaatg ttgggttccg taactggcat 2280
cccgttcccg caacgaccgg cgggaagaag tacggtggaa aaggtcacaa cggcggcgtc 2340
tgggaatgga cttcaactac gctcacgaaa cacgatggat tcgaaccatc taagcactac 2400
ccaggattca cggcggattt ctttgacgga tgtcacaacg tgattttggg cgggtcatac 2460
gcattcatcc ctcgcatggc tgaaaggcgc agtttgcgca actggtacca gaggaactac 2520
ccgtatgctt ggacgggagg gcgaattgtt tacgatattt ga 2562
<211> 414
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 3
Met Leu Lys Tyr Phe Ser Phe Glu Pro Gly Tyr Ile Asn Leu Asn Asn
1 5 1015
Gly Ser Phe Gly Ser Leu Pro Lys Pro Val Ala Glu Phe Cys Val Gln
202530
His Ala Ala Arg Val Glu Ala Asn Pro Asp Arg Tyr His Arg Leu Glu
354045
Phe Ala Glu Leu Leu Thr Ala Ala Arg Ala Arg Val Ala Arg Leu Ile
505560
Gly Ala Asp Thr Asp Glu Cys Val Phe Val Pro Ser Thr Thr Ala Gly
65707580
Ile Asn Thr Ile Leu Arg Asn Ile Glu Trp Ser Ser Gly Asp Val Ile
859095
Ile Arg Phe Ser Thr Thr Tyr Gln Gly Val Glu Arg Val Ala Arg Tyr
100 105 110
Ile Ala Asp Thr His Ser Glu Val Thr Leu Ser Thr Val Glu Leu Thr
115 120 125
Phe Pro Ile Ser His Gln Glu Ile Val Lys Arg Phe Arg Glu His Val
130 135 140
Val Gly Val Lys Ala Ser Cys His Ser Ser Ser Arg Val Tyr Ala Ile
145 150 155 160
Ile Asp Ser Val Val Ser His Pro Gly Val Ala Leu Pro Trp Lys Gln
165 170 175
Met Thr Gln Ile Cys Arg Glu Glu Gly Val Cys Ser Ile Ile Asp Ala
180 185 190
Ala His Ser Ile Gly Gln Glu Met Asn Ile Asp Leu Ala Ser Ala Gln
195 200 205
Pro Asp Phe Trp Val Ser Asn Cys His Lys Trp Leu Phe Ala Lys Arg
210 215 220
Gly Ser Thr Val Leu Tyr Val Pro Lys Arg Asn Gln His Ile Ile Lys
225 230 235 240
Ser Ala Leu Val Thr Ser Trp Asp Tyr Pro Ser Pro Lys Glu Ser Ala
245 250 255
Pro Ala Ala Tyr Asp Thr Gly Phe Val Leu Gln His Glu Trp Pro Gly
260 265 270
Ala Ile Asp Phe Ser Pro Tyr Leu Ser Val Cys Ala Ala Met Asp Phe
275 280 285
Arg Glu Lys Ile Gly Gly Glu Gln Ala Ile Asn Asp Tyr Cys His Lys
290 295 300
Leu Ala Leu Glu Gly Gly Arg Lys Leu Ala Ser Ala Leu Gly Gly Cys
305 310 315 320
Val Leu Asp Glu Ser Pro Ser Ala Glu Leu Thr Leu Asn Met Val Asn
325 330 335
Val Gln Leu Pro Leu Pro Pro Thr Thr Ser Lys Glu Leu Gln Gly Lys
340 345 350
Ile Asp Ser Phe Leu Glu Tyr Glu Leu Leu His Lys His Asn Val Ser
355 360 365
Ser Val His Phe Tyr His Gly Gly Ser Trp Trp Val Arg Cys Ser Ala
370 375 380
Gln Ile Trp Asn Glu Val Ser Asp Phe Glu Tyr Leu Gly Lys Ala Leu
385 390 395 400
Ser Asn Ala Cys Gln Glu Ala Arg Ser Lys Phe Phe Ala Asp
405 410
<211> 1245
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 4
atgctaaagt acttttcatt tgagcccgga tacatcaacc tcaacaatgg ttcatttggg 60
tccctcccca agccggtggc tgagttttgt gtgcaacatg ctgccagggt tgaagcgaat 120
ccggacagat atcatcgact cgagtttgcg gagctcctca cagctgcgcg cgctcgtgtt 180
gccagactga tcggagcaga cacggacgaa tgcgtgtttg ttccgagcac cactgcaggc 240
atcaacacaa tattacggaa cattgaatgg agttcaggcg atgtaatcat tagattctca 300
acaacttatc aaggggtgga acgggttgct agatacatag ccgacactca ttccgaagtg 360
acgctatcaa ccgtggagtt gacatttccc atctctcatc aagagattgt aaaacgcttc 420
cgagaacacg ttgttggggt caaggcaagc tgtcacagct catcgagggt ctacgccata 480
atcgactcgg tcgtatcgca cccgggtgtt gctttgccct ggaaacagat gacccaaatc 540
tgtcgcgaag agggcgtctg cagcattatt gacgcagccc actccattgg ccaggagatg 600
aacatagatc tggcatctgc gcagccagac ttttgggtat cgaactgcca caaatggctt 660
ttcgccaaac gtggttctac cgttttatat gtgccgaagc ggaaccaaca catcatcaag 720
tctgcgctgg taacatcctg ggactatccc tcccctaagg agtctgctcc tgctgcgtac 780
gatactggtt tcgttctcca acatgaatgg ccgggggcga tcgacttttc gccttacctg 840
agtgtctgtg ctgcgatgga cttccgcgag aagatcggtg gcgaacaggc gatcaacgac 900
tattgccaca agctggcact ggagggcggc cggaaattag cttctgctct tggaggttgt 960
gtcctcgacg agtcaccttc tgcggaattg acactcaaca tggtgaatgt acaactaccg 1020
cttcctccga caacatcgaa agagctccag ggaaagatcg atagctttct ggaatacgag 1080
ctgttacata aacacaacgt atcctccgtg cacttctacc acggtggttc gtggtgggtg 1140
cgctgcagtg ctcaaatttg gaacgaggtt tctgacttcg agtaccttgg caaagctctg 1200
agcaacgcgt gtcaagaggc tagatcgaag ttcttcgcgg actag 1245
<211> 855
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 5
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0