

IPC分类号 : C12N9/24I,C12N15/81I,C12N15/62I,C12P33/00I








专利摘要
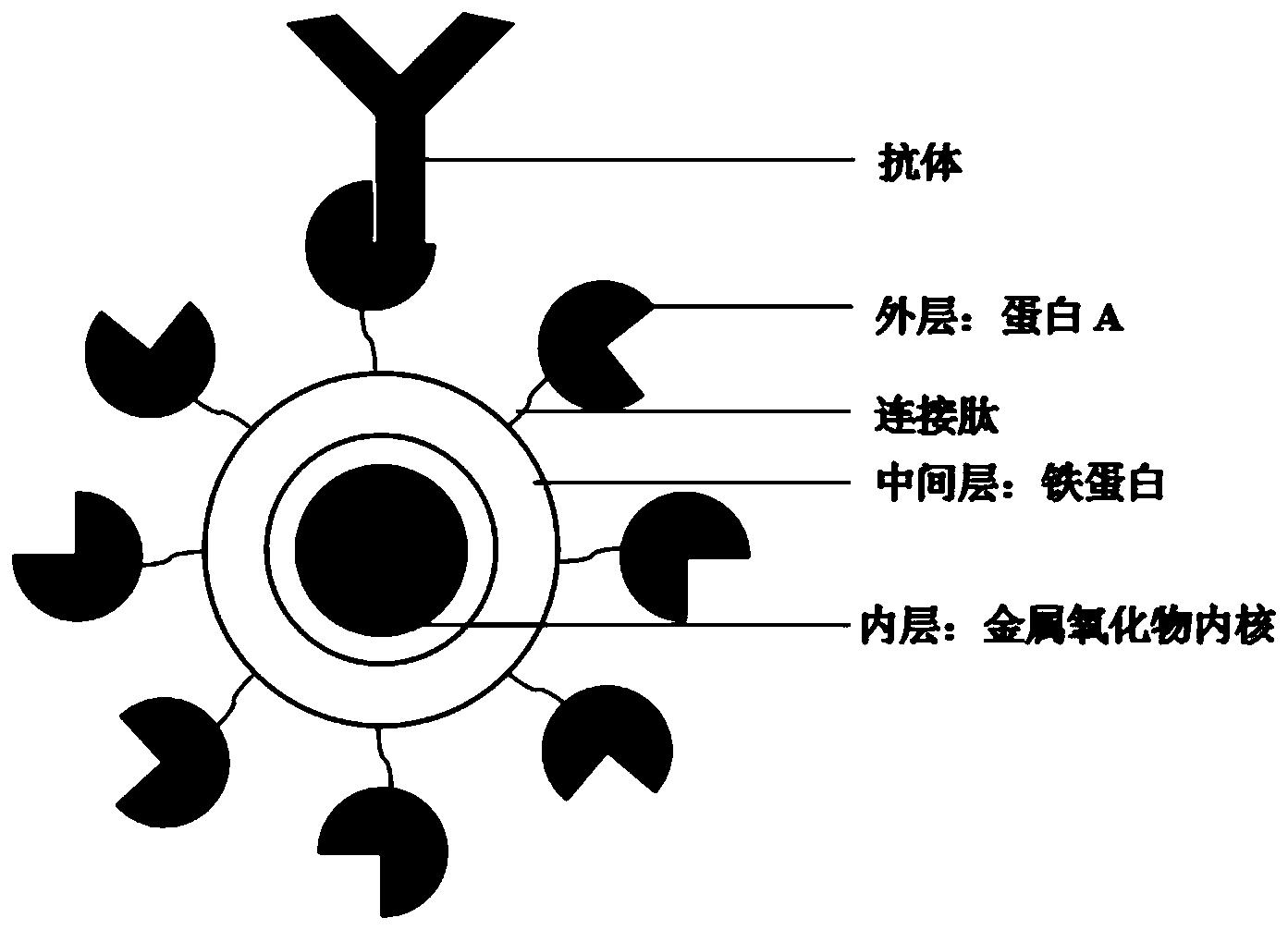
本发明涉及一种提高毕赤酵母表面展示β‑葡萄糖醛酸苷酶活性的方法。该方法包括:(1)将来自于酿酒酵母基因组的Agα1蛋白的编码基因与来源于米曲霉β‑葡萄糖醛酸酶(PGUS)的编码基因融合,导入到毕赤酵母细胞中,得到通过Agα1表面展示PGUS的重组酵母细胞;(2)将来自于酿酒酵母基因组的Pir1蛋白的编码基因与PGUS的编码基因融合,导入(1)获得的重组毕赤酵母细胞中,得到Agα1与Pir1两种锚定蛋白共同展示PGUS的重组酵母菌株。实验证明,这种方法能够显著增加毕赤酵母表面展示PGUS蛋白的量,且大幅提高了PGUS的稳定性。该方法解决了通过酶解法转化甘草酸为甘草次酸过程中存在的GUS不稳定与低效率的问题,同时对优化酵母表面展示效率的研究提供了方法指导。
权利要求
1.一种提高β-葡萄糖醛酸苷酶活性的毕赤酵母,其特征在于,包含:从N端至C端依次装载有β-葡萄糖醛酸苷酶基因pgus序列、linker基因序列和锚定蛋白基因pir1序列的pGAPZα表达载体;
所述β-葡萄糖醛酸苷酶基因pgus序列如SEQ ID NO.3所示;
所述linker基因序列为AAGCTTAGCGGCAGCGGTAGC;
所述锚定蛋白基因pir1序列如SEQ ID NO.2所示。
2.一种提高β-葡萄糖醛酸苷酶活性的毕赤酵母,其特征在于,包含:从N端至C端依次装载有β-葡萄糖醛酸苷酶基因pgus序列、linker基因序列和锚定蛋白基因pir1序列的pGAPZα表达载体,以及,从N端至C端依次装载有β-葡萄糖醛酸苷酶基因pgus序列、linker基因序列和锚定蛋白基因agα1序列的pGAPZα表达载体;
所述β-葡萄糖醛酸苷酶基因pgus序列如SEQ ID NO.3所示;
所述linker基因序列为AAGCTTAGCGGCAGCGGTAGC;
所述锚定蛋白基因pir1序列如SEQ ID NO.2所示;
所述锚定蛋白基因agα1序列如SEQ ID NO.1所示。
说明书
技术领域:
本发明涉及一种提高毕赤酵母表面展示β-葡萄糖醛酸苷酶酶活的方法,属于合成生物学范围。
背景技术:
甘草次酸是一种来自于甘草根部的齐墩果酸型三萜类化合物,分子式为C30H46O4,相对分子质量470.69。作为甘草酸的主要活性成分,甘草次酸具有适用于医学治疗和保健的各种生理功能和药物活性,例如抗肿瘤、抗炎、抗病毒、抗溃疡、抗过敏、抗氧化和肾上腺皮质激素样等,被广泛应用于在临床医药领域。甘草次酸可以通过作为甘草中最主要活性成分的甘草酸水解掉两分子葡萄糖醛酸基获得,该水解过程可以通过β-葡萄糖醛酸苷酶催化实现,目前甘草次酸的生产方法由酶的产生、酶的纯化、甘草酸的转化三个部分构成。该生产过程由于耗时高成本高,同时伴随着游离的β-葡萄糖醛酸酶不稳定的问题,因此严重影响了甘草酸转化在食品和医药行业的应用。
为了克服这些障碍,本发明中提出了酵母表面展示β-葡萄糖醛酸酶的方案。目前,酵母细胞表面展示已广泛应用于各种领域,如提高蛋白质的产量和热稳定性,cDNA文库筛选等。酵母表面展示除了利用酵母本身在发酵方面的巨大优势外,表面展示酶在保持其高效专一及温和的酶催化反应特性的同时,又克服了游离酶的不足之处,呈现存储稳定性高,分离回收容易,可被多次重复使用,连续操作可控,工艺简便等优点。因此,构建β-葡萄糖醛酸酶的表面展示菌株进行甘草酸的转化具有很大的实际意义。同时,使用蛋白表达和分泌过程最具成本效益的真核蛋白质表达系统巴斯德毕赤酵母作为表面展示宿主菌使工程菌的工业价值得到进一步提高。
虽然表面展示具有诸多优点,但仍存在展示蛋白量不高的问题,表面展示效率仍是制约其使用的关键,因此如何提高毕赤酵母展示的蛋白量仍是一个急需解决的问题,对酵母表面展示效率进行创新性的优化无论对科研还是对实际应用都有巨大意义。
发明内容:
1、本发明的一个目的是提供一种在毕赤酵母表面展示来源米曲霉β-葡萄糖醛酸酶(PGUS)的方法。
本发明提供的利用毕赤酵母表面展示PGUS的方法,具体可包括:
(1)将PGUS的基因(Genbank注册序列号为EU095019)与Pir1锚定蛋白的基因通过linker融合,构建到载体上获得质粒1,该融合蛋白记为融合蛋白1;将PGUS的基因与Agα1锚定蛋白的基因通过linker融合,构建到载体上获得质粒2,该融合蛋白记为融合蛋白2;
(2)质粒1插入毕赤酵母基因组,获得表面展示菌株,命名为PGUS-Pir;质粒2插入毕赤酵母基因组,获得表面展示菌株,命名为PGUS-Agα;
(3)重组毕赤酵母细胞PGUS-Pir表达了PGUS与Pir1的融合蛋白,该融合蛋白位于毕赤酵母细胞壁上;重组毕赤酵母细胞PGUS-Agα表达了PGUS与Agα的融合蛋白,该融合蛋白同样位于毕赤酵母细胞壁上。所述两个融合蛋白PGUS都位于N端;
上述方法所采用的受体酵母细胞具体为毕赤酵母GS115。
上述方法利用pGAPZα载体为带有博来霉素抗性的质粒,可以在大肠杆菌与毕赤酵母之间穿梭,融合蛋白的编码基因插入该质粒GAP启动子后的多克隆位点区,质粒构建完成后与毕赤酵母基因组进行同源重组,将目的基因插入到毕赤酵母基因组GAP位点。
上述方法中用于PGUS与锚定蛋白融合的linker为柔性linker,以GlySerlinker(GSGSGS)为基础进行改造的linker(KLSGSGS)。
上述方法中,融合蛋白进行表面展示所采用的信号肽为α-factor信号肽,位于融合蛋白基因的前端。
上述方法中,融合蛋白编码的基因中,PGUS基因位于5’端,锚定蛋白基因位于3’端。
上述方法中,融合蛋白C端都含有6X组氨酸的编码基因序列作为标签序列。
2、本发明的第二个目的是提供一种提高PGUS表面展示效率的方法。
本发明提供的提高PGUS表面展示效率的方法,具体可包括:
(1)利用重组毕赤酵母细胞PGUS-Agα制备感受态细胞;
(2)对质粒1的基础上进行插入HIS4筛选标记的改造,获得质粒3;
(3)质粒3插入PGUS-Agα基因组,获得既表达融合蛋白1又表达融合蛋白2的表面展示菌株,将命名为PP-1。
3、本发明的第三个目的是利用上述表面展示PGUS菌株在转化甘草酸生成甘草次酸中的应用。
在实际实施过程中,上述步骤包含如下具体工艺:
(1)培养上述构建的工程酵母72h,离心收集菌体;
(2)菌体加入到50ml 2g/L的甘草酸中,分别在40℃、50℃、60℃条件下反应5min;
(3)反应完成后通过测定甘草酸的转化效率确定最适的反应温度。
(4)离心1mL发酵液,取菌体,菌体加入到1ml 2g/L的甘草酸中,在确定的最佳温度下反应5min;
(5)反应完毕后,离心回收菌体,水洗两次后重新加入到50mL甘草酸中,确定重复利用率。
所述反应在pH 5~6下进行。
所述反应产物通过HPLC检测。
本发明具有催化效率高,反应条件温和,高效,绿色安全等优点。
附图说明:
图1本发明实施例5中展示酶在30-90℃的相对活性。
图2本发明实施例5中展示酶在pH 3.0-11.0的相对活性。
图3本发明实施例5中展示酶在70℃热孵育下不同时间的剩余活性。
图4本发明实施例6中展示酶在不同温度下的转化率。
图5本发明实施例7中展示酶的重复利用率。
具体实施方式:
下面结合实施例,对本发明的具体实施方式进一步详细描述。以下实施例用于说明本发明,但不用来限制本发明的范围。
本发明所述生物材料来源的一般性说明:
1、引物合成:本发明中所使用引物均由金唯智生物技术有限公司合成制备。
2、实验中所使用的T4DNA连接酶购自New England BIOlabS公司;FastPfu高保真聚合酶购自全式金公司;限制性内切酶均购自Thermo Fisher公司;使用的DNA胶回收试剂盒及质粒小提试剂盒及酵母基因组试剂盒均购自天根公司。
实验例1:质粒构建
A:锚定蛋白基因的获得
培养酿酒酵母INVSc1菌株24h后取5mL菌液进行基因组的提取。利用酿酒酵母INVSc1基因组上通过通过引物通过PCR扩增出目的基因。扩增体系为:FastPfu Buffer(2×)5.0μL、dNTP(2.5mM)2μL、模板0.5μL、引物(5μM)各2μL、FastPfu高保真DNA聚合酶0.5μL、补充双蒸水至25μL。扩增条件为95℃预变性3分钟;95℃变性20秒、60℃退火30秒、72℃延伸1分钟(30个循环);72℃延伸10分钟。通过天根琼脂糖凝胶DNA回收试剂盒回收得到agα1,pir1基因片段。上游引物Agα-F/Pir-F引入HindIII酶切位点,下游引物Agα-R/Pir-R引入NotI酶切位点(下划线标出)。
Agα-F CCCAAGCTTAGCGGCAGCGGTAGCGCTAGCGCCAAAAGC
Agα-R ATAAGAATGCGGCCGCAGACGAAAACAGCATGGACGATAAG
Pir-F CCCAAGCTTAGCGGCAGCGGTAGCGCCGCTGCTATC
Pir-R ATAAGAATGCGGCCGCAGAAGTTAAAGTTGTGGCTTGG
B:PGUS基因的获得
根据β-葡萄糖醛酸酶PGUS基因(Genbank注册序列号为EU095019)序列设计引物,PGUS的编码的基因序列为模板扩增,上游引入EcoRI酶切位点,下游引入HindIII酶切位点,回收得到带有酶切位点的pgus基因片段。
PGUS-F CCGGAATTCATGCTCAAACCGCAGCAAACTAC
PGUS-R CCCAAGCTTTCACTGGAAGGTCTTCCCTCCCTCAGCAGT
C:质粒组装
限制性核酸内切酶EcoRI和NotI双酶切pGAPZα质粒;内切酶EcoRI和HindIII双酶切回收的agα1、pir1片段;内切酶HindIII和NotI双酶切pgus片段。
酶切体系:酶A 2μL和酶B 2μL,10x digest buffer 5μL,DNA片段30μL,用双蒸水补足50μL的酶切体系,酶切条件为37℃、2h,随后80℃反应20分钟使酶失活。通过天根琼脂糖凝胶DNA回收试剂盒回收得到pGAPZα、agα1、pgus,通过T4连接酶连接;将酶切回收后的产物pGAPZα、pir1、pgus控制连接片段的摩尔比通过T4连接酶连接。两种连接产物热击后分别转化至E.coli TOP 10感受态细胞中,涂布于含有50mg/L博来霉素的固体LB培养基(蛋白胨10g/L,酵母提取物5g/L,氯化钠10g/L,琼脂糖20g/L)平板上,37℃培养。
采用菌落PCR和测序方法鉴定转化子。菌落PCR体系:模板LB平板单菌落,pGAPZα上下游通用引物各1μL,2xrTaq mix(宝生物公司)10μL,用双蒸水补足20μL。PCR条件:预变性94℃5min,变性94℃30s、退火60℃30s、延伸72℃1min 30s,循环30次,72℃10min,4℃保存。经菌落PCR验证含有目标条带的转化子送DNA测序,确定质粒构建成功。测序结果表明质粒构建正确,agα1序列为序列表中SEQ ID NO:1、pir1核苷酸序列为序列表中SEQ ID NO:2、pgus核苷酸序列为序列表中SEQ ID NO:3。
实施例2:构建表面展示PGUS的工程菌
重组载体pGAPZ-Pir-PGUS及pGAPZ-Agα-PGUS构建完成后,使用限制性内切酶AvrII线性化质粒并回收,制作毕赤酵母GS115感受态细胞(现做现用),然后将回收的线性质粒通过电转化法转入新鲜制备的GS115感受态中,涂布在含有1/2000Zeocin的YPDZ平板上培养,直至出现明显可见的酵母单菌落。挑取单菌落在YPDZ平板上划线保存。从保存的YPDZ划线平板上随机挑取重组菌,每个挑取3个平行对照组,接种到装有5mL的YPDZ液体培养基中的试管中培养约24h。取2mL菌液提取酵母基因组,然后用相应的上下游引物进行PCR验证扩增目的条带,扩增出目的条带的重组菌初步断定为阳性菌。最终得到的两种表面展示PGUS的阳性菌株,分别命名为PGUS-Pir、PGUS-Agα。
实施例3:酵母免疫荧光验证
为了验证PGUS被成功展示到毕赤酵母细胞壁表面,分别接种PGUS-Pir、PGUS-Agα和空白GS115菌株,培养36h后,按1:1000的稀释率稀释作为一抗的His-Tag的小鼠IgG。收集酵母细胞并重悬于含有1mg/mL牛血清白蛋白(BSA)和一抗的PBS缓冲液中,并在冰上孵育30min,每5min轻微震荡一次防止酵母沉淀。然后洗涤细胞并重悬于含有第二抗体(1:1000稀释)红藻中提取的天然橙红色荧光色素PE的抗小鼠抗体,在PBS缓冲液中避光放置30min,偶尔翻转,用PBS(无BSA)洗涤两次后,将细胞重悬于40μL PBS缓冲液中,并且将5-10μL悬浮液点样在载玻片上以在UV光下显微观察。荧光显微镜观察结果到明显的红色荧光,说明PE荧光二抗已经牢固锚定了6*His一抗,一抗紧密结合了酵母细胞壁上的PGUS蛋白。PGUS-Pir、PGUS-Agα的荧光效果类似,酵母边缘明显更亮。
实施例4:展示酶比酶活测定
(1)β-葡萄糖醛酸苷酶以甘草酸为底物催化产物检测:
反相高效色谱法建立定量测定生物转化甘草酸分析方法。仪器主要参数:岛津LC-10A,色谱柱:Shim-pack,VP-ODS,检测器:SPD-10AVP氘灯检测器,检测波长:254nm,流动相A为甲醇,流动相B为6‰的醋酸溶液,进样量:10μL,流速:1mL/min,柱温箱:40℃,工作站:LCsolution。
取100mL展示工程菌菌体反应液,离心,弃上清,加入400mL 50mM pH 5.0乙酸-乙酸钠缓冲液配制的2g/L甘草酸溶液。40℃条件下反应30min后,加入10μL 2M NaOH溶液使反应终止,然后取100μL反应液,加入到900μL甲醇中混匀,用0.22μm水系滤膜过滤后,HPLC检测底物甘草酸和产物甘草次酸的含量。
酶活力定义:在上述反应条件下,每分钟催化水解1μmol甘草酸所需的细胞干重为一个活力单位(U)。
以甘草酸为底物,PGUS-Pir的比酶活28.89U/g,是PGUS-P的35倍。
实施例5:展示酶酶学性质的测定
(1)PGUS展示酶的最适催化温度测定
最适催化反应温度:最适催化反应温度是将不同展示酶与pH为5、浓度为0.79mM的p-NPG分别在不同温度(30-80℃)下反应,检测酶活,检测不同展示酶的最适催化温度。
图1显示PGUS-Pir的最适温度为60℃,且最适温度范围很大,75℃仍保有90%的酶活。PGUS-Agα,其最适温度为65℃,在70℃后酶活迅速下降。对比游离PGUS(最适温度为40℃),展示酶的最适温度整体有了约20度的移动。
(2)展示酶最适pH的测量
最适催化反应pH:分别将不同的展示酶与不同pH(3-11)配制的0.79mM p-NPG进行反应,按2.2.12的方法检测酶活,检测酶的最适反应pH。使用的不同pH(3-11)缓冲液为:0.1mol/L甘氨酸-盐酸缓冲液(pH 1.0-3.0);0.1mol/L醋酸缓冲液(pH 4.0-5.0);0.1mol/L磷酸钾缓冲液(pH 6.0-7.0);0.1mol/L Tris-HCI缓冲液(pH 8.0-10.0);0.1mol/L甘氨酸-氢氧化钠缓冲液(pH 11.0-12.0)。
接种PGUS-Pir,PGUS-Agα。配置展示酵母反应液改用不同pH(3-11)的缓冲液配制,通过pNPG检测酶活。实验室前期对PGUS-P的最适pH进行过摸索,游离酶的最适pH都在5.0附近。结果见图2,表面展示酶的最适pH整体已经发生了很大的偏移,定位在了pH 7.0附近。
(3)PGUS展示酶的热稳定性测定
温度稳定性:将展示酶放于70℃水浴中保温180min,分别在不同时间间隔取样,按pNPG法测定酶活力,以零时刻的酶活作为100%参照算出不同保温时间下的剩余酶活力,确定为β-葡萄糖醛酸苷酶的温度稳定性。
温度70℃孵育时,展示酶的稳定性开始受到强烈影响,总体来说残余活性随着孵育时间逐渐减少。结果见图3,游离的PGUS半衰期为52min,进行表面展示后,PGUS-Agα半衰期为180min,PGUS-Pir在热处理30min时酶活反而有所提高,在超过1h的热处理后酶活几乎没有损失,继续延长PGUS-Pir的热处理时间后,结果显示其热处理5h后PGUS-Pir仍有70%以上酶活,最终在360min时达到半衰期,热稳定性对比游离酶提高了6.92倍。
实施例6:使用双锚定蛋白提高展示效率
由于使用pGAPZa进行第一次重组后,工程菌已携带有博来霉素抗性,因此第二个质粒必须含有其他的抗性基因或缺陷型基因。GS115的常用缺陷型为His,可以载体上携带his4基因,染色体上His4位点与载体上his4基因之间发生单十字交换事件后会在His4位点形成插入。
采用在质粒pGAPZa上添加His4标记的方法。从pPIC9K载体上通过His4-F/His4-R扩增下his4基因,大小为2535bp,切胶回收。通过pGAP-his-F/pGAP-his-R扩增pGAPZ-Pir-PGUS质粒,切胶回收,通过Gbison组装的方法,在pGAPZ-Pir-PGUS载体上插入His4标记。Gibsonassembly反应体系:片段与载体共2.5μL,Gibson Assembly master mix 6.5μL,Taqligase 1.0μL。
pGAP-his-F TAACCAATTCTGACCTTCGTTTGTGCGGATCCCCCAC
pGAP-his-R GCATTCTGACATCCTCTTGATTAGAATCTAGCAAGACCGGTCT
His4-F GTCTTGCTAGATTCTAATCAAGAGGATGTCAGAATGCCATTTGC
His4-R CGCACAAACGAAGGTCAGAATTGGTTAATTGGTTGTAACACTGG
获得pGAPZ-His4-Pir-PGUS载体,使用AvrII作为质粒的线性化位点,培养工程菌PGUS-Agα,将其制备感受态细胞,线性化的pGAPZ-His4-Pir-PGUS点交换到PGUS-Agα工程菌的基因组GAP区。涂布于缺少组氨酸、含有2%葡萄糖的基础培养基SD-HIS平板,培养48小时,通过菌落PCR筛选阳性转化子,获得的阳性菌株命名为PP-1。
接种PP-1、PGUS-Pir、PGUS-Agα三个工程菌,培养72h,由于毕赤酵母的高密度发酵特性,取30μ发酵液稀释100倍测OD600,然后根据OD值稀释发酵液到等OD。等OD等体积的菌体通过Tris-HCL缓冲液制备反应液,使用甘草酸测定酶活。
结果见图4,以甘草酸为底物,等量的PP-1酶活是PGUS-Pir的144%。
实施例7:全细胞高温催化甘草酸的水解工艺
表面显示使PGUS反应性能得到了显著提高,本节设计了一种新的反应过程,充分利用展示PGUS,以期提高反应效率。
使用50mL的2g/L甘草酸作为底物,在40℃-60℃的温度下进行测试。测量PGUS转化率、PGUS-Pir转化率和PP-1转化率,其中游离的PGUS由于酶活较低,在酶浓度不足的情况下存在底物抑制现象,因此将其酶浓度相比于展示酵母提高40倍进行反应。结果如图4所示,PP-1在60℃转化率达到了92.1%,而在40℃只有73.6%的转化率。游离的PGUS得益于高酶量,在40℃获得较高转化率(88.7%),但在高温条件下转化率急剧下降。
从转化效率分析,60℃更适合作为展示酵母进行甘草酸转换的温度。然后对PP-1与PGUS-Pir进行重复利用率的测定,进行如下操作:
(1)离心1mL发酵液,取菌体;
(2)菌体加入到1ml 2g/L的甘草酸中,60℃反应5min;
(3)反应完毕后,离心回收菌体,水洗两次后重新加入到1mL甘草酸中;
重复十次次上述操作,测定每次反应甘草酸转化量,以第一次反应为100%。结果如图5所示,十次重复使用PP-1后仍存在75%的酶活力。
基于展示酵母的高重复利用率,完成分批补料工艺的设计,该工艺1mL OD600=6的发酵液对应的菌体在60℃下转化甘草酸的空间转化率为146.7g·L-1d-1,且通过回收菌体进行多次重复利用。
序列表
<110> 北京理工大学
<120> 一种提高毕赤酵母表面展示β-葡萄糖醛酸苷酶活性的方法
<160> 3
<170> SIPOSequenceListing 1.0
<210> 1
<211> 966
<212> DNA
<213> Saccharomyces cerevisiae
<400> 1
gctagcgcca aaagctcttt tatctcaacc actactactg atttaacaag tataaacact 60
agtgcgtatt ccactggatc catttccaca gtagaaacag gcaatcgaac tacatcagaa 120
gtgatcagtc atgtggtgac taccagcaca aaactgtctc caactgctac taccagcctg 180
acaattgcac aaaccagtat ctattctact gactcaaata tcacagtagg aacagatatt 240
cacaccacat cagaagtgat tagtgatgtg gaaaccatta gcagagaaac agcttcgacc 300
gttgtagccg ctccaacctc aacaactgga tggacaggcg ctatgaatac ttacatcccg 360
caatttacat cctcttcttt cgcaacaatc aacagcacac caataatctc ttcatcagca 420
gtatttgaaa cctcagatgc ttcaattgtc aatgtgcaca ctgaaaatat cacgaatact 480
gctgctgttc catctgaaga gcccactttt gtaaatgcca cgagaaactc cttaaattcc 540
ttctgcagca gcaaacagcc atccagtccc tcatcttata cgtcttcccc actcgtatcg 600
tccctctccg taagcaaaac attactaagc accagtttta cgccttctgt gccaacatct 660
aatacatata tcaaaacgga aaatacgggt tactttgagc acacggcttt gacaacatct 720
tcagttggcc ttaattcttt tagtgaaaca gcactctcat ctcagggaac gaaaattgac 780
acctttttag tgtcatcctt gatcgcatat ccttcttctg catcaggaag ccaattgtcc 840
ggtatccaac agaatttcac atcaacttct ctcatgattt caacctatga aggtaaagcg 900
tctatatttt tctcagctga gctcggttcg atcatttttc tgcttttgtc gtacctgcta 960
ttctaa 966
<210> 2
<211> 471
<212> DNA
<213> Saccharomyces cerevisiae
<400> 2
gccgctgcta tctctcaaat tggtgacggt caaatccaag ccactaccaa aaccactgct 60
gctgctgttt ctcaaattgg tgacggtcaa atccaagcca ctactaaaac caaagctgct 120
gctgtctctc aaattggtga cggccaaatc caagccacca ccaagactac ctcagctaag 180
actaccgctg cagccgtctc ccaaattggt gacggtcaaa ttcaagccac tactaaaacc 240
aaagctgctg ctgtctctca aattggtgac ggtcaaatcc aagccactac caaaacaact 300
gctgcagctg tctctcaaat tggtgacggt caaatccaag ccactactaa aaccactgct 360
gctgctgttt ctcaaattgg tgacggtcaa atccaagcca ccaccaatac tactgttgct 420
ccagtctccc aaatcactga tggccaaatc caagccacaa ctttaacttc t 471
<210> 3
<211> 1815
<212> DNA
<213> Aspergillus oryzae
<400> 3
atgctcaaac cgcagcaaac tactacccga gacctcatct cgctcgacgg gctatggaaa 60
tttgccctag catccgacga caacaatacg caaccatgga caagccaact aaaaacgtcc 120
ctggaatgcc cggtcccagc atcctacaat gacatattcg cagacagtaa gatccacgat 180
cacgtcggat gggtgtacta ccagcgcgac gtgattgtgc caaagggctg gtccgaagaa 240
cggtatcttg tgcgctgtga agccgcaaca caccatggcc ggatctacgt caatggaaat 300
ctggtcgcgg accatgtggg cggttacacg ccgtttgagg cagatattac cgatctggtt 360
gctgccggag aacaattccg cctgaccatt gcggttgaca atgaactcac gtatcagacg 420
atcccacctg gaaaggtgga gattctcgag gcgacgggca agaaggtgca gacttatcag 480
catgattttt acaattatgc tgggctggct cggtcggtgt ggctgtattc tgtgccacag 540
cagcacattc aggatatcac tgttcggacg gatgtgcagg gcaccaccgg gctgattgac 600
tacaatgttg tggccagtac gacacaagga accattcagg ttgccgtgat agatgaggat 660
ggcacaaccg tagcgacaag ctcaggatcg aacgggacta tccatatacc atccgttcac 720
ctttggcaac ctggcgccgc gtatctgtac caactccatg ccagcatcat agactcttcc 780
aaaaagacta ttgatacata taagctagca acgggcatcc ggacggtcaa agtacagggt 840
actcagttcc tcataaacga caagcccttc tacttcaccg ggttcggtaa acacgaggac 900
actaatatcc gcggcaaagg ccacgacgac gcctacatgg tgcacgactt tcagctgctg 960
cactggatgg gggccaactc cttccgaacc tcacactacc catacgccga ggaggtcatg 1020
gaatatgccg atcgacaggg catagtcgtg atagatgaaa cacccgccgt cggtctagca 1080
ttctccatcg gagcgggagc ccagacgtct aacccgccgg ctacattcag cccagatcgc 1140
ataaacaaca agactcgcga agctcacgct caagctatcc gtgagcttat ccaccgcgat 1200
aaaaaccatc ccagcgtggt catgtggtcc atcgctaacg agcctgcctc taacgaggac 1260
ggcgcccgcg aatacttcgc acctctgccc aagctggctc gtcagcttga cccgacgcga 1320
cctgtcactt tcgccaatgt gggtcttgct acctataagg cagaccgtat tgcagacctc 1380
tttgatgtgc tttgcttaaa ccggtatttt ggttggtaca cgcaaacggc tgagctcgac 1440
gaggcagaag ctgcgctgga agaagaactt cgcggatgga ccgagaagta tgacaaaccg 1500
attgtcatga cggagtacgg agcagacacg gtggcgggct tgcactcggt catggttact 1560
ccctggagcg aggagttcca ggtcgagatg ctggatatgt atcatcgggt ctttgatcgg 1620
ttcgaggcca tggcaggtga acaagtctgg aatttcgcgg actttcaaac tgcggttggt 1680
gttagtcgag tggacgggaa caagaaaggt gtttttacta gggatcggaa gccaaaggct 1740
gctgctcatc tattgagaaa gaggtggacg aacttgcata atggtactgc tgagggaggg 1800
aagaccttcc agtga 1815
一种提高毕赤酵母表面展示β-葡萄糖醛酸苷酶活性的方法专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。















动态评分
0.0