

IPC分类号 : C12N9/10I,C12N15/54I,C12N15/74I,C12P19/42I,C12N13/00I,C12N1/20I,C12R1/41N







专利摘要
本发明公开一种甲硫氨酸合成酶突变体、突变基因及其在制备维生素B12中的应用。在苜蓿中华根瘤菌中过表达的所述甲硫氨酸合成酶基因和突变基因的基因工程菌,其生产维生素B12的能力得到了大幅提高,并且对发酵培养时的生物量几乎没有任何影响,具有较大的应用推广价值。
权利要求
1.一种甲硫氨酸合成酶的突变体,其特征在于,其氨基酸序列在如SEQ ID NO:4所示氨基酸序列存在下述突变:第136位氨基酸突变为L,或者第136位氨基酸突变为L,并且第245位氨基酸突变为S。
2.编码如权利要求1所述甲硫氨酸合成酶的突变体的编码基因。
3.如权利要求2所述的编码基因,其特征在于,其核苷酸序列如SEQ ID NO:2或SEQ IDNO:3所示。
4.甲硫氨酸合成酶编码基因在制备维生素B12中的应用,其特征在于,所述甲硫氨酸合成酶编码基因编码具有SEQ ID NO:5或SEQ ID NO:6所示氨基酸序列的多肽。
5.如权利要求4所述的应用,其特征在于,通过含有所述编码基因的表达载体导入苜蓿中华根瘤菌中进行过表达。
6.如权利要求5所述的应用,其特征在于,所述苜蓿中华根瘤菌具有CGMCC NO.9638保藏号的菌株。
7.如权利要求4所述的应用,其特征在于,所述甲硫氨酸合成酶编码基因具有SEQ IDNO:2或SEQ ID NO:3所示核苷酸序列。
8.如权利要求5至7任一项所述的应用,其特征在于,所述导入的编码基因位于质粒或染色体中。
说明书
技术领域
本发明属于生物技术领域,具体涉及到甲硫氨酸合成酶突变体、突变基因及其在制备维生素B12中的应用。
背景技术
维生素 B12 (VB12)在药物和食品工业中具有广泛的应用,其又叫钴胺素,属于咕啉类化合物,是唯一含金属元素的维生素类化合物,是B族维生素发现最晚的大分子有机化合物。根据咕啉环上方的配基(R基团)种类不同,维生素B12可分为:羟基钴胺素,脱氧腺苷钴胺素和甲基钴胺素。维生素B12参与了大量的生化反应过程包括DNA的合成和调控、脂肪酸的合成、氨基酸的代谢及能力的产生。
由于维生素B12分子结构复杂,化学法人工合成需要消耗大量的人力与物力,而且合成周期长。在合成过程中对操作人员的要求过高,导致不能大量的生产。微生物发酵法目前是生产维生素B12的最廉价的方法,能够大量生产并推广其使用。
目前,国内外针对维生素B12生产菌生物合成维生素B12的研究主要集中在发酵过程优化,主要涉及培养基包括碳氮源和金属离子的优化、甜菜碱的添加、鱼藤酮的添加,工艺条件包括pH和供氧的控制等。有文献报道通过在脱氮假单胞菌基因组上表达单拷贝的透明颤菌vgb 基因来增加细胞生产维生素B12的能力(程立芳等, 不同来源的尿卟啉原 III转甲基酶在脱氮假单胞菌 中的表达及其对生产维生素 B12 的影响. 工业微生物,2017,第 47 卷 第 3期)。
S-腺苷甲硫氨酸(SAM)是广泛存在于生物体内的一种重要中间体,参与多种类型的生化反应,主要包括转甲基、转硫和转氨丙基。在细胞内,SAM是由S-腺苷甲硫氨酸合成酶(S-adenosy-L-methionine synthetase, SAMS, EC2.5.1.6)催化三磷酸腺苷(ATP)和底物L-甲硫氨酸(L-Met)合成的。5-甲基四氢叶酸-高半胱氨酸甲基转移酶,又叫甲硫氨酸合成酶(5-methyltetrahydrofolate--homocysteine methyltransferase, EC:2.1.1.13),是催化N5一甲基四氢叶酸的甲基转移到半胱氨酸上生成甲硫氨酸的酶(Han G et al,Metabolic engineering of Corynebacterium glutamicum ATCC13032 to produce S-adenosyl-L-methionine. Enzyme Microb Technol. 2016 Feb;83:14-21. doi:10.1016/j.enzmictec.2015.11.001.)。 SAM是维生素B12合成的重要甲基供体,而甲硫氨酸的供给对SAM的合成至关重要。目前,尚未有甲硫氨酸合成酶编码基因metH用于维生素B12合成的报道。
发明内容
本发明人前期筛选出了一株高产维生素B12的苜蓿中华根瘤菌CGMCC NO.9638菌株(CN104342390A),可发酵生产维生素B12。通过对其进行诱变,获得产维生素B12能力提高的诱变菌株,再挖掘分析其中能够增加产维生素B12的能力相关的基因,本发明其中甲硫氨酸合成酶基因及其突变基因对于苜蓿中华根瘤菌产维生素B12的能力有重要影响,因而做了进一步研究,最终完成本发明。
本发明提供一种甲硫氨酸合成酶的突变体,其氨基酸序列在如SEQ ID NO:4所示氨基酸序列存在下述突变:第136位氨基酸突变为L,或者第136位氨基酸突变为L,并且第245位氨基酸突变为S。更具体地,其氨基酸序列如SEQ ID NO:5或SEQ ID NO:6所示。
本发明进一步提供编码如上述甲硫氨酸合成酶的突变体的编码基因。更具体地,其核苷酸序列如SEQ ID NO:2或SEQ ID NO:3所示。
本发明尤其提供甲硫氨酸合成酶编码基因在制备维生素B12中的应用。
在一个具体实施方式中,其是通过含有所述编码基因的表达载体导入苜蓿中华根瘤菌中进行过表达。进一步地,所述导入的编码基因位于质粒或染色体中。
优选地,所述苜蓿中华根瘤菌具有CGMCC NO.9638保藏号的菌株。
其中,所述甲硫氨酸合成酶编码基因优选编码具有SEQ ID NO:4、SEQ ID NO:5或SEQ ID NO:6所示氨基酸序列的多肽。
更优选地,所述甲硫氨酸合成酶编码基因具有SEQ ID NO:1、SEQ ID NO:2或SEQID NO:3所示核苷酸序列。
经对比研究证实,本发明中,过表达甲硫氨酸合成酶基因和突变基因的工程菌生物安全,对发酵培养的生物量几乎没有任何影响,尤其可以有效的提高苜蓿中华根瘤菌生产维生素B12的能力。经对比实验,数据表明其中对于原始基因在苜蓿中华根瘤菌中过表达,可以提高产维生素B12的能力达11.5%,而突变基因在苜蓿中华根瘤菌中过表达能够进一步提高产维生素B12的能力(可达20%以上)。
附图说明
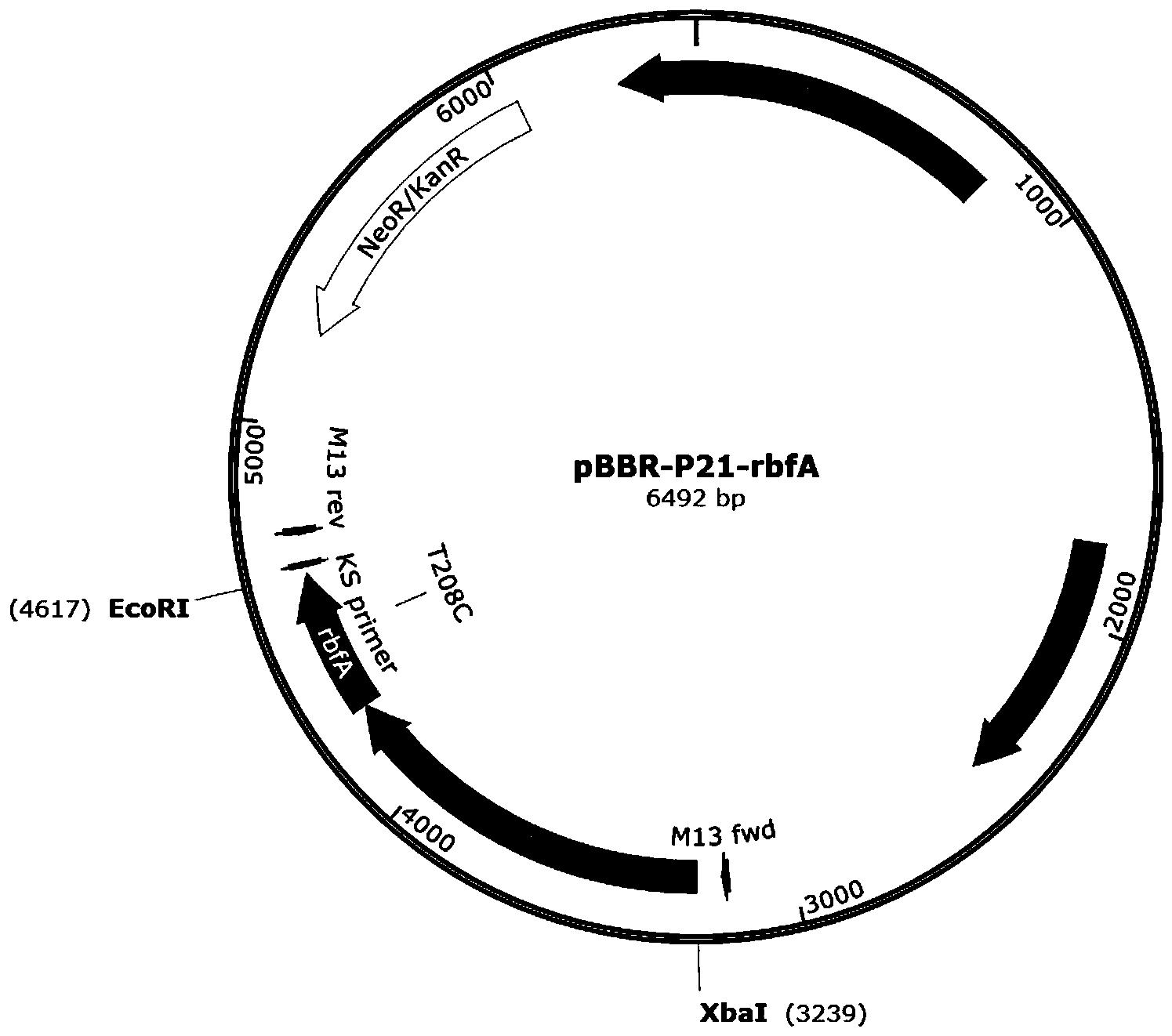
图1:质粒载体pBBR-P21-metH的图谱。
图2:不同苜蓿中华根瘤菌菌株发酵144h后的VB12产量。
图3:不同苜蓿中华根瘤菌菌株发酵144h后的生物量。
图4:维生素B12的标准曲线图。
具体实施方式
本发明的以下实施例和附图仅说明实现本发明的具体实施方案,这些方案和附图不可以理解为对本发明的限制,任何在不脱离本发明的原理和实质的情况下所做的任何改变,均落在本发明的保护范围之内。
本实施例中所用到的实验技术与实验方法,如无特殊说明均为常规技术方法。本实施例中所使用的材料、试剂等,如无特殊说明,均可通过正规商业渠道获得。
培养基配方:
LB 培养基(g/L):氯化钠 10 ,胰蛋白胨10 ,酵母提取物5,固体培养基加琼脂粉15。
种子培养基(g/L):蔗糖40,玉米浆20,甜菜碱5,(NH4)2SO4 1,(NH4)2HPO4 2,MnSO4·H2O 0.8,CoCl2·6H2O 0.02,MgO 0.3,DMBI 0.01,ZnSO4·7H2O 0.01,CaCO3 1.5,pH通过NaOH控制在7.0~7.4。
发酵培养基(g/L):蔗糖80,玉米浆30,甜菜碱15,(NH4)2SO4 2,MgSO41.5, K2HPO40.75,CoCl2·6H2O 0.14, DMBI 0.075,ZnSO4·7H2O 0.08,CaCO3 1,pH通过NaOH控制在7.0~7.4。
检测方法
(1) 样品预处理
取1 mL发酵液,加入8%的亚硝酸钠溶液和冰醋酸各0.25mL摇匀,置于95~100℃水浴中30-40 min;待冷却至室温,在10000转离心1分钟,上层清液通过0.22μm膜(⌀= 0.22µm)过滤器过滤至上样瓶中,再加20μL 2%的NaCN (w / v)至1mL的上清液中。亚硝酸钠溶液和冰醋酸的加入量可随发酵液的量进行相应调整。
(2)标准品的制备
配置梯度维生素B12标准品(20 mg/L,50 mg/L,100mg/L,150 mg/L)。
(3)HPLC检测条件
C18-250A柱(Agilent,4.6 mmid 9×250 mm,5µm)。流动相为70%有机相(乙腈)和 30%无机相(乙酸钠水溶液),吸收波长为361 nm,柱温为35℃,流速0.8 mL/min,进样量20 µL。
(4)维生素B12标准曲线的绘制
将不同浓度的标准品按上述条件进行HPLC检测,绘制峰面积A-VB12浓度标准曲线。以测得的峰面积A为纵坐标,维生素B12质量浓度C(mg/L)记为横坐标,绘制维生素B12标准曲线。见图4,得回归方程y=19.846x-80.857,R2=0. 999,吸收度与质量浓度呈良好的线性关系。液相结束后根据维生素B12标准曲线计算样品产量。
实施例1:常压室温等离子体(ARTP)诱变时间的确定、突变体库的构建及高产菌种的获得
(1)致死率的测定
为了获得一个比较广的突变体库,我们对底盘细胞苜蓿中华根瘤菌CGMCCNO.9638进行了常压室温等离子体(ARTP)诱变。首先,对等离子诱变条件下CGMCCNO.9638的致死率进行测定。将种子用LB培养基培养至对数中期的CGMCCNO.9638细胞(OD600=1) 用0.85% NaCl溶液冲洗两遍,然后用0.85% NaCl溶液稀释成108个细胞/mL的悬液。取10uL悬液均匀的涂布在铁片上,进行ARTP诱变,诱变时间分别为0 s、5 s、10 s、15 s、20 s、25 s,每个诱变时间点2个重复,每个时间点的涂板3个重复。诱变后将带有细胞的铁片置于1 mL无菌水中漩涡震荡洗下细胞,将细胞悬液10倍梯度稀释至10-3,取稀释后的细胞悬液100 μL涂LB培养基平板,30℃培养72 h后统计菌落数。根据下列公式计算不同诱变时间下的致死率,以诱变时间为横坐标,不同诱变时间下的致死率为纵坐标,绘制致死率曲线。致死率的计算公式为:致死率(%)=[(诱变0s菌落数-诱变Ns菌落数)/诱变0s菌落数]×100%,其中N=5、10、15、20、25。
从表1中可知,诱变10s时的致死率为82.2%,诱变15s时的致死率达到95%以上,致死率为80%-90%时,诱变后发生正突变的概率最高,因此选择10s作为最终构建突变体库的诱变时间。
(2)突变体库的构建及筛选
取浓度为108个/mL的细胞悬液10 uL涂布于铁片上进行ARTP诱变,诱变时间为10 s,共对3个铁片上的细胞进行诱变处理,诱变结束后,将这3个铁片上的细胞在含有1mL LB培养基的1.5 mL离心管中涡旋振荡重悬。将细胞悬液10倍梯度稀释至10-3,取稀释后的细胞悬液100 μL涂LB培养基平板,30℃培养72h,长出270个单菌落。
(3)突变株96深孔板发酵初筛
分别挑取平板上的所有单菌落接种于含有500uL的种子培养基的96深孔板中(每个孔板上包含6株对照菌株CGMCC NO.9638),30℃、800rpm、80%湿度振荡培养36h后,按10%(v/v)接种量转接含有450uL发酵培养基的96深孔板中,30℃、800rpm、80%湿度条件下振荡培养120h后检测维生素B12产量。
(4)突变株96深孔板发酵复筛
将步骤(3)中产量排前30的株菌的单菌落(包含一个对照菌株CGMCC NO.9638)接种于含有500uL种子培养基的96深孔板中,30℃、800rpm、80%湿度振荡培养36h后,按10%(v/v)接种量转接含有450uL发酵培养基的96深孔板中(每株菌3个平行),30℃、800rpm、80%湿度条件下振荡培养120h后检测维生素B12产量。
重复步骤(3)和(4)三次,最后获得一株高产维生素B12的菌种SM*,96孔板中产量从50 mg/L提高到80 mg/L,是底盘菌株产量的1.6倍。
实施例2:突变菌株与出发菌株进行比较基因组分析
将菌种SM*和出发菌株CGMCC NO.9638送金唯智生物科技有限公司进行全基因组测序。通过比较两个菌株的全基因组序列发现甲硫氨酸合成酶编码基因metH发生了点突变,其第406位核苷酸上含有突变,其所述突变是用C置换了T,在第733位核苷酸上也含有突变,其所述突变是用A置换了G。为了验证突变位点对维生素B12产量的影响,将突变前和突变后的metH基因在出发菌种CGMCC NO.9638中进行了过量表达。
实施例3:质粒载体的构建
1、pBBR-P21-metH的构建:
分别利用表2的引物对P21-XbaI-F和P21-R,以Ensifer adhaerens Casida A(黏着剑菌)基因组为模板,通过PCR 扩增,引入XbaI酶切位点,得到启动子P21片段,经电泳验证,用限制性内切酶DpnI(NEB公司)在37℃处理30min,核酸电泳胶回收后,得到纯化的P21片段。P21启动子序列如SEQ ID No.7所示,并且记载在专利申请号为201910929398.X的专利文件中。
分别利用表2的引物对metH-F和metH-XhoI-R,以苜蓿中华根瘤菌CGMCCNO.9638基因组为模板,通过PCR 扩增,引入XhoI酶切位点,得到metH片段,经电泳验证,DpnI酶法处理,电泳胶回收后,得到纯化的metH片段。metH基因序列如SEQ ID No.1所示,其编码的氨基酸序列如SEQ ID No.4所示。
然后,利用引物对P21-XbaI-F和metH-XhoI-R,以纯化的P21片段和metH片段为模板,通过融合PCR,得到P21-metH片段(含XbaI和XhoI酶切位点),经电泳验证,电泳胶回收后,得到纯化的P21-metH片段。
将上述纯化的P21-metH片段和pBBR1MCS2质粒分别用XbaI和XhoI进行双酶切,将P21-metH片段的双酶切产物与pBBR1MCS2质粒双酶切的产物经过T4连接酶4℃过夜连接。将连接产物转化入大肠杆菌DH5α中,涂布于含有50mg/L卡那霉素的LB固体平板上,培养16h后进行菌落PCR检测,送金唯智测序,测序正确后,将得到的阳性菌命名为E.coli/pBBR-P21-metH。用质粒试剂盒提取质粒pBBR-P21-metH备用,质粒图谱如图1所示。
、pBBR-P21-metH(T406C)
以质粒pBBR-P21-metH为模板,用引物对 T406C-F/ T406C-R进行反向PCR扩增,得到大小约7.1kb的片段DpnI酶法处理,电泳胶回收后,得到纯化的产物。取30ng纯化后的产物加入2μL 10*T4连接酶缓冲液(NEB公司)、1μL T4多核苷酸激酶(NEB公司),补充蒸馏水至20μL,37℃反应30 min,加入1μL T4连接酶(NEB公司),室温反应2 h得到连接产物。将连接产物转化入大肠杆菌DH5α中,涂布于含有50mg/L卡那霉素的LB固体平板上,培养16h后进行菌落PCR检测,送金唯智测序,测序正确后,将得到的阳性菌命名为E.coli/pBBR-P21-metH(T406C),其中涉及metH基因的核苷酸序列如SEQ ID NO. 2所示,氨基酸序列如SEQ ID NO.5所示。用质粒试剂盒提取质粒pBBR-P21-metH(T406C)备用。
、pBBR-P21-metH(T406C,G733A)
以质粒pBBR-P21-metH(T406C)为模板,用引物对G733A-F/ G733A-R进行反向PCR扩增,得到大小约7.1kb的片段DpnI酶法处理,电泳胶回收后,得到纯化的产物。取30ng纯化后的产物加入2μL 10*T4连接酶缓冲液(NEB公司)、1μL T4多核苷酸激酶(NEB公司),补充蒸馏水至20μL,37℃反应30 min,加入1μL T4连接酶(NEB公司),室温反应2 h得到连接产物。将连接产物转化入大肠杆菌DH5α中,涂布于含有50mg/L卡那霉素的LB固体平板上,培养16h后进行菌落PCR检测,送金唯智测序,测序正确后,将得到的阳性菌命名为E.coli/pBBR-P21-metH(T406C,G733A), 其中涉及metH基因的核苷酸序列如SEQ ID NO. 3所示,氨基酸序列如SEQ ID NO. 6所示。用质粒试剂盒提取质粒pBBR-P21-metH(T406C,G733A)备用。
实施例4:含质粒载体菌株的构建
将实施例3中的4个质粒pBBR1MCS2、pBBR-P21-metH、pBBR-P21-metH(T406C)和pBBR-P21-metH(T406C,G733A)按照如下方法转入苜蓿中华根瘤菌CGMCCNO.9638中:
(1)接种新活化的苜蓿中华根瘤菌CGMCCNO.9638、大肠杆菌(含有相应的质粒)以及辅助载体MT616,并分别在30℃和37℃的培养箱中振荡培养到OD值为1.0左右;
(2)无菌条件下分别将苜蓿中华根瘤菌CGMCCNO.9638的菌液、MT616和大肠杆菌的菌液各500 μL转移至1.5mL无菌EP管中并在4 ℃,12,000 rpm条件下离心1 min。
(3)在无菌条件下弃掉上清液,并用1mL 0.85%的无菌生理盐水对沉淀进行悬浮。
(4)再次在4℃,12,000 rpm条件下离心1min,并在无菌条件下去除上清。
(5)分别采用500μL新鲜的LB液体培养基对受体细胞、大肠杆菌和MT616的沉淀进行悬浮。
(6)分别取各2μL的三种菌液,滴在不添加抗性的LB固体培养基的同一位置,并小心将其混匀。分别将单一组分的菌液以及两两之间混合的菌液进行点样,用来作为试验对照组。
(7)待菌液自然风干后,在37℃培养箱中倒置培养约1天,至单菌落长出。
(8)挑取不同的单菌落在含有相应抗生素的平板上进行划线,将平板倒置于30℃培养箱中进行培养,直到菌落长出。同时对照组也需挑取不同的单菌落在含有相应抗生素的平板上进行划线。
(9)从抗性平板上挑取长出的克隆,进行菌落PCR验证。得到阳性苜蓿中华根瘤菌:SM/pBBR (对照菌)、SM/pBBR-P21-metH(简写为SM1)、SM/ pBBR-P21-metH(T406C)(简写为SM2)、SM/ pBBR-P21-metH(T406C,G733A)(简写为SM3)。
实施例5:不同菌株评价
1、苜蓿中华根瘤菌的培养条件:
将对照菌、SM1、SM2和SM3菌株在无菌条件下用接种针在含100mg/L卡那霉素的LB固体培养基上划线,30℃恒温静置48 h培养,获得单菌落。用接种针挑取单菌落于装有5 mL含有100mg/L卡那霉素的LB液体培养基的试管中,30℃,200 rpm培养36h。将种子培养基按照10%比例接种至含有100mg/L卡那霉素的30mL发酵培养基中(250ml摇瓶)。30℃震荡(220 r/min)培养144 h后收集菌体并检测产量。摇瓶发酵每个实验做3个平行。
、不同苜蓿中华根瘤菌菌株产VB12能力的比较
SM1、SM2和SM3三个菌株与对照菌相比生产维生素B12的能力均得到提高。结果见表3和图2。
其中SM1提高11.5%,SM2提高20.7%,SM3提高 24.1%。结果说明本发明中过量表达甲硫氨酸合成酶编码基因metH的苜蓿中华根瘤菌菌株生产维生素B12的能力得到显著提高。四个菌株生物量变化不大,说明metH基因的过表达未对菌体的生长造成影响(见表3和图3)。
序列表
<110>中国科学院天津工业生物技术研究所
<120>甲硫氨酸合成酶突变体、突变基因及其在制备维生素B12中的应用
<160>15
<170> PatentIn Version 3.1
<210>1
<211> 1017
<212>DNA
<213>Sinorhizobium meliloti
<400> 1
atgaccgcag cctccaatgc cctctccgct ctcatcgccg aaaaaggcgt tcttctcgcc 60
gacggcgcca ccggcacatc gctcttcgcc atgggcctcg aagcaggcga agcgccggaa 120
ctctggaacg aaacgaagcc ggagaacatc accaagctgc accaggattt cgtcgacgcc 180
ggcgccgaca tcatcctcac caacaccttt ggcggcaccc gtcaccggct gaaactgcac 240
caggccgacg accgcgtgca tgcgctgaac aagcgcgccg ccgagatcgc ccgtgcggtt 300
gccgacaagg catcgcgcaa ggtcatcacc gccggctccg tcggcccgac cggcgaactg 360
ctgatccccc ttggcgcact ttcctatgaa gatgcagtta ccgccttcgt cgagcagatc 420
gaggggctga aggcgggcgg cgccgaagtc gcctggatcg agaccatgtc ctcgccggac 480
gagatccgtg cggccgccga agcggccgcc aaggtcggcc ttccctacgt ctataccggc 540
tcgttcgata ccgccggcaa gacgatgatg gggctgcacc cgaaggatat ccatggcgtt 600
gccaccgagt tcggcgacgg cccggtcgcc gttggcgcca attgcggcgt aggtgcttcc 660
gacattctct cctcgctgct cgacatgacg gaggcagcac ccgaggcaac ggtgatcgtc 720
aagggcaatt gcggcatccc ggaattccgc ggctcggaga ttcactattc cggcacgccg 780
ccgctgatgg ccgaatatgt gcgccttgcg gccgatgccg gcgccaagat catcggcggc 840
tgctgcggca ccacctgcga ccacctcgcc gccatgcgcg tcgcactcga cgaacatgtg 900
cgccgcgaac gcccgacgct cgacctgatc atcgaaaaga tcggcccgct gcgcaacaag 960
accgccaacg aaacggcggc cgcgcctgcc cgcgagcgcc gcagccgccg cgcctga 1017
<210>2
<211>1017
<212>DNA
<213>Sinorhizobium meliloti
<400> 2
atgaccgcag cctccaatgc cctctccgct ctcatcgccg aaaaaggcgt tcttctcgcc 60
gacggcgcca ccggcacatc gctcttcgcc atgggcctcg aagcaggcga agcgccggaa 120
ctctggaacg aaacgaagcc ggagaacatc accaagctgc accaggattt cgtcgacgcc 180
ggcgccgaca tcatcctcac caacaccttt ggcggcaccc gtcaccggct gaaactgcac 240
caggccgacg accgcgtgca tgcgctgaac aagcgcgccg ccgagatcgc ccgtgcggtt 300
gccgacaagg catcgcgcaa ggtcatcacc gccggctccg tcggcccgac cggcgaactg 360
ctgatccccc ttggcgcact ttcctatgaa gatgcagtta ccgccctcgt cgagcagatc 420
gaggggctga aggcgggcgg cgccgaagtc gcctggatcg agaccatgtc ctcgccggac 480
gagatccgtg cggccgccga agcggccgcc aaggtcggcc ttccctacgt ctataccggc 540
tcgttcgata ccgccggcaa gacgatgatg gggctgcacc cgaaggatat ccatggcgtt 600
gccaccgagt tcggcgacgg cccggtcgcc gttggcgcca attgcggcgt aggtgcttcc 660
gacattctct cctcgctgct cgacatgacg gaggcagcac ccgaggcaac ggtgatcgtc 720
aagggcaatt gcggcatccc ggaattccgc ggctcggaga ttcactattc cggcacgccg 780
ccgctgatgg ccgaatatgt gcgccttgcg gccgatgccg gcgccaagat catcggcggc 840
tgctgcggca ccacctgcga ccacctcgcc gccatgcgcg tcgcactcga cgaacatgtg 900
cgccgcgaac gcccgacgct cgacctgatc atcgaaaaga tcggcccgct gcgcaacaag 960
accgccaacg aaacggcggc cgcgcctgcc cgcgagcgcc gcagccgccg cgcctga 1017
<210>3
<211>1017
<212>DNA
<213>Sinorhizobium meliloti
<400> 3
atgaccgcag cctccaatgc cctctccgct ctcatcgccg aaaaaggcgt tcttctcgcc 60
gacggcgcca ccggcacatc gctcttcgcc atgggcctcg aagcaggcga agcgccggaa 120
ctctggaacg aaacgaagcc ggagaacatc accaagctgc accaggattt cgtcgacgcc 180
ggcgccgaca tcatcctcac caacaccttt ggcggcaccc gtcaccggct gaaactgcac 240
caggccgacg accgcgtgca tgcgctgaac aagcgcgccg ccgagatcgc ccgtgcggtt 300
gccgacaagg catcgcgcaa ggtcatcacc gccggctccg tcggcccgac cggcgaactg 360
ctgatccccc ttggcgcact ttcctatgaa gatgcagtta ccgccctcgt cgagcagatc 420
gaggggctga aggcgggcgg cgccgaagtc gcctggatcg agaccatgtc ctcgccggac 480
gagatccgtg cggccgccga agcggccgcc aaggtcggcc ttccctacgt ctataccggc 540
tcgttcgata ccgccggcaa gacgatgatg gggctgcacc cgaaggatat ccatggcgtt 600
gccaccgagt tcggcgacgg cccggtcgcc gttggcgcca attgcggcgt aggtgcttcc 660
gacattctct cctcgctgct cgacatgacg gaggcagcac ccgaggcaac ggtgatcgtc 720
aagggcaatt gcagcatccc ggaattccgc ggctcggaga ttcactattc cggcacgccg 780
ccgctgatgg ccgaatatgt gcgccttgcg gccgatgccg gcgccaagat catcggcggc 840
tgctgcggca ccacctgcga ccacctcgcc gccatgcgcg tcgcactcga cgaacatgtg 900
cgccgcgaac gcccgacgct cgacctgatc atcgaaaaga tcggcccgct gcgcaacaag 960
accgccaacg aaacggcggc cgcgcctgcc cgcgagcgcc gcagccgccg cgcctga 1017
<210>4
<211>338
<212>PRT
<213>Sinorhizobium meliloti
<400> 4
MTAASNALSA LIAEKGVLLA DGATGTSLFA MGLEAGEAPE LWNETKPENI TKLHQDFVDA 60
GADIILTNTF GGTRHRLKLH QADDRVHALN KRAAEIARAV ADKASRKVIT AGSVGPTGEL 120
LIPLGALSYE DAVTAFVEQI EGLKAGGAEV AWIETMSSPD EIRAAAEAAA KVGLPYVYTG 180
SFDTAGKTMM GLHPKDIHGV ATEFGDGPVA VGANCGVGAS DILSSLLDMT EAAPEATVIV 240
KGNCGIPEFR GSEIHYSGTP PLMAEYVRLA ADAGAKIIGG CCGTTCDHLA AMRVALDEHV 300
RRERPTLDLI IEKIGPLRNK TANETAAAPA RERRSRRA 338
<210>5
<211>338
<212>PRT
<213>Sinorhizobium meliloti
<400>5
MTAASNALSA LIAEKGVLLA DGATGTSLFA MGLEAGEAPE LWNETKPENI TKLHQDFVDA 60
GADIILTNTF GGTRHRLKLH QADDRVHALN KRAAEIARAV ADKASRKVIT AGSVGPTGEL 120
LIPLGALSYE DAVTALVEQI EGLKAGGAEV AWIETMSSPD EIRAAAEAAA KVGLPYVYTG 180
SFDTAGKTMM GLHPKDIHGV ATEFGDGPVA VGANCGVGAS DILSSLLDMT EAAPEATVIV 240
KGNCGIPEFR GSEIHYSGTP PLMAEYVRLA ADAGAKIIGG CCGTTCDHLA AMRVALDEHV 300
RRERPTLDLI IEKIGPLRNK TANETAAAPA RERRSRRA 338
<210>6
<211>338
<212>PRT
<213>Sinorhizobium meliloti
<400> 6
MTAASNALSA LIAEKGVLLA DGATGTSLFA MGLEAGEAPE LWNETKPENI TKLHQDFVDA 60
GADIILTNTF GGTRHRLKLH QADDRVHALN KRAAEIARAV ADKASRKVIT AGSVGPTGEL 120
LIPLGALSYE DAVTALVEQI EGLKAGGAEV AWIETMSSPD EIRAAAEAAA KVGLPYVYTG 180
SFDTAGKTMM GLHPKDIHGV ATEFGDGPVA VGANCGVGAS DILSSLLDMT EAAPEATVIV 240
KGNCSIPEFR GSEIHYSGTP PLMAEYVRLA ADAGAKIIGG CCGTTCDHLA AMRVALDEHV 300
RRERPTLDLI IEKIGPLRNK TANETAAAPA RERRSRRA 338
<210>7
<211> 1000
<212>DNA
<213>Ensifer adhaerens Casida A
<400> 7
caaacagacc gggatatgcg ggtattcttc cgccgcgccg aggatgaggt ggcgcaggaa 60
cgcgtcaccg gcataggagc gggcgccacg gcttgcctga aggatgaccg ggctgtcggt 120
cgcatcggcg gcgcgcatga cggcctgaat gtattccaga ttgttcacat tgaacgccgg 180
cagcgcgtaa tcgttctccg ccgcatggtc gagcagttgc cgcaatgtga tcaatgccat 240
tcgctatctc cctttggata ctcggtgcaa cctatgcggc gcaccacaaa aacaatccgg 300
ccgttgaacc gcacgaaatg catcgatggc aaagtcgatg gccggctttt tcgtgcggcg 360
tgacggcgcg cgcgaattgg tcgcgcccac cgaagtcagg cgcacaatag ttcatcgaag 420
tggtttgaca accgggcaaa aggcaggttg ccagaggtcg aaactcgctt caatcgattt 480
tactgtggac tggatgcaac accttcagtg tgaagtgttt tcactttctg gtggtgcctg 540
agaggagggg gagtcgaggg cagtggatgc aaccattggg cgctgatttt gtctgttaca 600
ccatcgtggt ggatgccctg tcggaaacag tctgtcgaca ggaggtgaac gtcccgcaag 660
aagattgcgg caacgcccct ctttctttgc gcagattacg taaactgccg ctaaaattca 720
caaactttgc atcgcggatg attcgaggct caatccggcc gacaaaaagc gcggacctaa 780
aacgttgcag tagatttcgc aaaaatgccc tgttcacgtc atatgcccgt cgcaaaggcg 840
acgaaaagaa tcgcaaacaa aatacaacct atgggatagg ccgattcccc tcctatagat 900
aaagatgcag acagccgcag aatccgcctt gcgttcgcga acgatttgcg cttctctcct 960
gcgatcacaa acccaaaaca aggggaagga gagaaacaaa 1000
<210>8
<211>32
<212>DNA
<213>人工序列
<400> 8
ctagtctaga caaacagacc gggatatgcg gg 32
<210>9
<211> 40
<212>DNA
<213>人工序列
<400>9
gcattggagg ctgcggtcat tttgtttctc tccttcccct 40
<210>10
<211> 40
<212>DNA
<213>人工序列
<400> 10
aggggaagga gagaaacaaa atgaccgcag cctccaatgc 40
<210>11
<211>29
<212>DNA
<213>人工序列
<400> 11
ccgctcgagt caggcgcggc ggctgcggc 29
<210>12
<211>41
<212>DNA
<213>人工序列
<400> 12
atgaagatgc agttaccgcc ctcgtcgagc agatcgaggg g 41
<210>13
<211> 41
<212>DNA
<213>人工序列
<400> 13
cccctcgatc tgctcgacga gggcggtaac tgcatcttca t 41
<210>14
<211>41
<212>DNA
<213>人工序列
<400> 14
tgatcgtcaa gggcaattgc agcatcccgg aattccgcgg c 41
<210>15
<211> 41
<212>DNA
<213>人工序列
<400> 15
gccgcggaat tccgggatgc tgcaattgcc cttgacgatc a 41
甲硫氨酸合成酶突变体、突变基因及其在制备维生素B12中的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。


















动态评分
0.0