

IPC分类号 : C12P9/00,C12P13/00,C07F9/10,C12R1/38,C12R1/39,C12R1/40









专利摘要
本发明涉及用磷脂酶A2(phospholipaseA2)对从假单胞菌属(Pseudomonassp.)微生物提取的磷脂进行处理而生产溶血磷脂酰乙醇胺(lysophosphatidylethanolamine)18:1的方法,本发明的溶血磷脂酰乙醇胺18:1可以用于增加植物体的免疫反应的疫苗材料以及用作增进水果储藏性质的组合物,且无抑制植物体生长的副作用,进而能够用于食品、药品、化妆品和农业用途,因此在相关产业中非常有益。
权利要求
1.一种生产溶血磷脂酰乙醇胺的方法,其中,包括:
(a)培养假单胞菌属(Pseudomonas sp.)微生物后收获所述微生物并从所述微生物提取磷脂的步骤;
(b)用磷脂酶A
(c)将所述(b)步骤中生成的溶血磷脂酰乙醇胺进行分离和纯化的步骤;
其中,所述假单胞菌属微生物为淤泥假单胞菌(Pseudomonas caeni)、荧光假单胞菌(Pseudomonas fluorescens)、恶臭假单胞菌(Pseudomonas putida)或丁香假单胞菌(Pseudomonas syringae)。
2.根据权利要求1所述的方法,其特征在于,所述溶血磷脂酰乙醇胺为溶血磷脂酰乙醇胺18:1。
3.根据权利要求1所述的方法,其特征在于,所述磷脂为磷脂酰乙醇胺。
4.根据权利要求3所述的方法,其特征在于,所述磷脂酰乙醇胺的1位酰基链为18:1。
说明书
技术领域
本发明涉及由假单胞菌属(Pseudomonas sp.)微生物生产溶血磷脂酰乙醇胺18:1(lysophosphatidylethanolamine 18:1,以下称为LPE18:1)的方法,更详细而言,涉及用磷脂酶A2(phospholipase A2,以下称为PLA2)对从属于假单胞菌属的细菌微生物提取的磷脂进行处理而生产LPE18:1的方法。
背景技术
溶血磷脂酰乙醇胺(lysophosphatidylethanolamine),是作为细胞膜主要构成成分的磷脂(phospholipids)中的一种的磷脂酰乙醇胺(phosphatidyl ethanolamine)中的第二(2位,2nd)酰基链由于PLA2酶活性而脱离生成的,其在生物体内少量地存在。已知用溶血磷脂酰乙醇胺进行处理时,抑制植物体老化并促进水果的成熟(ripening)(美国授权专利第5110341号和美国授权专利第5126155号)。溶血磷脂酰乙醇胺主要用PLA2酶对从蛋黄或豆子中提取的磷脂酰乙醇胺进行处理而产生,组成成分大部分是第一(1位,1st)酰基链为16:0或18:0的LPE16:0和LPE18:0。在研究溶血磷脂酰乙醇胺以何种生物机制抑制老化的过程中明确了LPE18:1与LPE16:0或LPE18:0相比是具有更高生物功效的更优越的分子(superior molecule)(美国授权专利第6426105号)。目前为止,对于溶血磷脂酰乙醇胺制备方法的技术是将蛋黄或豆子用作原料,其成分大部分为LPE16:0和LPE18:0,几乎没有LPE18:1(韩国授权专利第0331932号)。为此,一直努力寻找生成LPE18:1的原料,但是没有找到。
本发明揭示了能够产生LPE18:1的原料在假单胞菌之类的微生物中,详细的研究方法和结果在下面叙述。如果病原菌侵入植物体,则植物体对一部分的病原菌发生抗病反应,但是其机制还不是很清楚。特别是关于初期抗病性诱导信号传导机制的信息不多。在此期间,已知当携带avrRpm1的丁香假单胞菌pv.番茄DC3000(Pseudomonas syringaepv.tomatoDC3000,以下称为Pst-avrRpm1)浸透宿主植物体拟南芥(Arabidopsisthaliana,Col-0 ecotype)的细胞外空间时,病原菌通过Ⅲ型分泌机制将Avr蛋白(avrRpm1)引入宿主的细胞质,如果宿主细胞具有识别avrRpm1蛋白的抗性蛋白即RPM1,则上述蛋白质之间的相互作用将引发水杨酸的增加、NPR1(PR基因1的非表达子(NON-EXPRESSER OF PR GENES 1))的活化以及PR(病程相关(PATHOGENESIS-RELATED))基因的表达等抗性基因诱导免疫反应。但是,连接这样的基因对基因(gene-for-gene)相互作用的上游阶段的分子、遗传学基础以及植物体的抗性基因诱导宿主免疫反应大部分还不明确。
本发明的发明人在这种还不明确的初期信号传导研究中明确了LPE18:1是诱导植物体的抗病性的信号传导因子。让人惊讶的是,明确了LPE18:1主要从侵入的病原菌(假单胞菌)中生成出来。总结的研究结果如下:如果非病原性假单胞菌(Pst-avrRpm1)侵入拟南芥植物体,则将avrRpm1蛋白注入细胞内,此时,植物体将该蛋白因子识别成基因与基因的相互作用(gene-for-gene interaction),立即使磷脂酶A2-α(PLA2α)蛋白表达。PLA2α蛋白向侵入的病原菌存在的细胞外分泌,分解病原菌的膜中存在的磷脂酰乙醇胺而生成LPE18:1作为主要成分。该LPE18:1发挥信号传导物质功能,向周围扩散,进入病原菌未侵入的相邻细胞内而诱导抗病性。
如上述研究结果所示,生成LPE18:1的原料存在于假单胞菌等微生物中。对此,本发明的发明人确认了大量培养假单胞菌后,提取脂质,用PLA2酶进行处理,从而大量生成LPE18:1。将这种自己制备的LPE18:1散布于植物体,并将其生物效果与现有的LPE16:0/LPE18:0混合液以及在市场上销售的高纯度的LPE18:1进行比较。由微生物假单胞菌自己制备的LPE大部分为LPE18:1和LPE16:0,并存在极少量的LPE16:1。自己制备的粗LPE18:1在效果方面优于LPE16:0/LPE18:0混合物,但是比纯LPE18:1低。另外,研究了生成LPE18:1的原料是否在假单胞菌以外的微生物中也能够得到。对于作为革兰氏阴性菌的大肠杆菌(E.coli)、作为革兰氏阳性菌的枯草芽孢杆菌(Bacillus subtilis)、柠檬节杆菌(Arthrobacter citresus)、作为酵母的酿酒酵母(Saccharomyces cerevisiae)、以及作为藻类的普通小球藻(Chlorella vulgaris),分离了磷脂,并进行分析,结果在其它微生物中用PLA2进行处理时,几乎不生成LPE18:1,只有在假单胞菌中生成。
另一方面,在韩国公开专利第2002-0086604号中公开了“增进植物的健康,保护植物免受生物和非生物性胁迫相关的损伤,促进植物从作为这种胁迫的结果的损伤中恢复的方法”,在韩国公开专利第1997-0001484号中公开了“用于使植物对病害免疫的方法及组合物”,但没有记载本发明的由假单胞菌属(Pseudomonas sp.)微生物生产溶血磷脂酰乙醇胺18:1的方法。
发明内容
技术问题
本发明是在上述情况下研究出来的,本发明的发明人确认了利用磷脂酶A2α(PLA2α)由侵入植物体的病原菌丁香假单胞菌的细胞膜磷脂产生了诱导植物免疫性的溶血磷脂酰乙醇胺18:1,基于此结果,大量培养假单胞菌(Pst)后,提取脂质,用PLA2酶进行处理,LPE18:1的大量产生被确证。另外,为了研究从假单胞菌以外的微生物中是否也能够获得LPE18:1的原料(含有18:1的第一酰基链的PE),磷脂从作为革兰氏阴性菌的大肠杆菌(E.coli)、作为革兰氏阳性菌的枯草芽孢杆菌(Bacillus subtilis)、柠檬节杆菌(Arthrobacter citresus)、作为酵母的酿酒酵母(Saccharomyces cerevisiae)、以及作为藻类的普通小球藻(Chlorella vulgaris)中分离,并用PLA2进行处理,结果确认了在其它微生物中包含LPE18:1的LPE的生成不多,只有在假单胞菌中明显生成。另外,为了确证在属于假单胞菌属的其它种中是否也能够大量产生LPE18:1,磷脂从从淤泥假单胞菌(Pseudomonas caeni)、荧光假单胞菌(Pseudomonas fluorescens)、恶臭假单胞菌(Pseudomonas putida)中被提取,结果确认了作为LPE的原材料的含有18:1的第一酰基链的PE的含量在假单胞菌的所有种中均相当多,用PLA2酶进行处理时,大量生成了LPE18:1。
因此,本发明确认了在大量培养假单胞菌微生物后,提取脂质,然后用由动物获得的PLA2进行处理,能够大量产生LPE18:1,从而完成了本发明。
解决问题的手段
为了解决上述问题,本发明提供用磷脂酶A2(phospholipase A2)对从假单胞菌属(Pseudomonas sp.)微生物提取的磷脂进行处理而产生溶血磷脂酰乙醇胺(lysophosphatidylethanolamine)18:1的方法。
发明效果
根据本发明,确证了利用磷脂酶A2α由侵入植物体的病原菌的细胞膜磷脂生成了诱导植物免疫性的粗溶血磷脂酰乙醇胺18:1,诱发植物体的局部性免疫反应。本发明的利用磷脂酶A2处理而产生的溶血磷脂酰乙醇胺18:1是来源于天然的物质,可以作为用于增加植物体的免疫反应的疫苗材料,以及用作增进水果储藏性质的组合物,且无抑制植物体生长的副作用,进而能够用于食品、药品、化妆品和农业用途,因此在相关产业中非常有益。
附图说明
图1是确认在pla2α变异植物体中局部性免疫力受到损伤的结果,A和B是观察Pst-avrRpm1渗入6天后,与野生型植物体不同地,在pla2α(敲除的,knockout)和pla2α-Ⅱ(敲低的,knockdown)变异植物体的整个植物体(A)和叶(B)中超出过敏性反应区域疾病的症状进展的照片。C是确认在野生型植物体和pla2α变异植物体的叶子中使Pst-avrRpm1(avrRpm1)、Pst-avrRpt2(avrRpt2)、Pst-avrRps4(avrRps4)和毒性的Pst(Pst)渗入15小时后的离子渗出的结果。NT:无处理对照组。D是Pst-avrRpm1渗入后第24小时,在植物体中确认PR1和PDF1.2基因的表达水平(*P<0.05)的结果。E是在野生型植物体、pla2α变异植物体和用天然启动子::PLA2α补偿的pla2α变异植物体(PLA2α/pla2α)中对Pst-avrRpm1和Pst感染细菌的生长进行分析的结果(*P<0.05,**P<0.01)。F是在野生型植物体、pla2α变异植物体中对Pst-avrRpt2和Pst-avrRps4感染细菌的生长进行分析的结果(**P<0.01)。
图2是确认暂时性的PLA2α的表达对LPE之类的脂质产物的生成带来的影响的结果,A是在接种了Pst-avrRpm1(avrRpm1)的野生型植物体中确认PLA2α表达的结果,B是在Pst-avrRpm1和Pst渗入1.5小时后在PLA2α启动子::GUS转化的拟南芥植物体中分析GUS活性的照片,C是在野生型植物体中对接种病原菌3小时后生成的脂质产物进行分析的结果,D是在pla2α变异植物体中对病原菌接种,根据病原菌接种和重组PLA2α蛋白处理的脂质产物进行分析的结果,E是在模拟(mock)处理叶和Pst-avrRpm1接种3小时后的野生型叶子中分析LPE种类的各水平的结果,F是分析了从将存活的Pst-avrRpm1、细菌脂质提取物及叶组织分别与PLA2α蛋白混合而得到悬浮液释放的LPE种类的结果,G是确认重组PLA2α蛋白的抗菌能力的结果,M表示用模拟(mock)进行处理的情形,P表示用PLA2α蛋白物进行处理的情形,PM表示用PLA2α蛋白与曼诺力得(manoalide)的混合物进行处理的情形,以及PMM表示用PLA2α蛋白与曼诺力得缓冲液(乙醇)的混合物进行处理的情形(*P<0.05,**P<0.01)。H是将进行了放射性标记的Pst-avrRpm1接种于野生型植物体的叶子后,3小时后确认放射性的程度的结果。
图3是确认LPE18:1导致的pla2α变异植物体的局部性免疫反应的恢复的结果,A是用LPE、LPC(溶血磷脂酰胆碱(lysophosphatidylcholine))和LPG(溶血磷脂酰甘油(lysophosphatidylglycerol))对感染了Pst-avrRpm1的pla2α变异植物体进行处理后观察表现型缺陷是否恢复的照片,B是用LPE或LPG对感染了Pst-avrRpm1的pla2α变异植物体进行处理后分析感染细菌的生长的结果,C是在经模拟(mock)处理或用LPE进行处理的野生型叶子中分析PR1基因、伤口相关基因的表达水平的结果,D是在野生型植物体和pla2α变异植物体中关于LPE处理导致的局部性免疫力的获得进行确认的结果,E是表示说明LPE导致的局部性免疫力获得的对毒性的Pst的抗病性增加的样子的照片,F是分析在体外中是否存在LPE自身对于Pst的抗菌活性的结果(*P<0.05,**P<0.01)。
图4是确认LPE导致的ICS1-依赖性水杨酸生物合成和NPR1活化的结果,A和C是确认基因表达水平的结果,B和D是分析杨酸水平的结果。E是对携带35S::NPR1-eGFP的转基因植物体进行模拟(mock)、LPE或水杨酸的处理,确认NPR1的从细胞质到核的位置变化的图片。
图5是确认PLA2α蛋白和由LPE介导的乙烯-依赖性PDF1.2的表达的结果,A和D是分析基因表达水平的变化的结果,B和E是分析乙烯合成水平的结果,以及C和F是分析茉莉酸的合成水平的结果(*P<0.05,**P<0.01)。
图6是表示在与正常相比不太成熟的叶子和短日照生长植物体叶子中pla2α变异植物体的缺陷不大的结果,确认了为了局部性免疫反应的诱导,PLA2α活性和PLA2α的质外体的分泌是重要的。A和B是接种Pst-avrRpm1后在pla2α变异植物体中区分生长条件(SD,9小时光照周期的短日照条件;LD,16小时光照周期的长日照条件)和叶子的成熟度程度来确认细菌生长的结果(*P<0.05,**P<0.01)。C是使缓冲液、Dsbc或PLA2α蛋白渗入pla2α变异植物体后确认ICS1和PR1基因的表达水平的结果,D是在模拟(mock)处理、PLA2α蛋白处理、以及将PLA2α蛋白和抑制剂(Mano)一同进行处理的组中分析PR1基因的表达的结果,E是用PLA2α抑制剂进行预处理并接种Pst-avrRpm1后,在野生型和pla2α变异植物体中分析细菌生长的结果,F是将具有PLA2α结构的载体转化(no S.P.PLA2α/PLA2α)进pla2α变异植物体后接种Pst-avrRpm1,并分析细菌生长的结果,上述PLA2α结构的载体由于没有信号肽,因此无法分泌到质外体而留在细胞质中。
图7是本发明的PLA2α蛋白和LPE信号传导物质在对于细菌侵入的免疫反应中起到何种作用的模式图。
图8是表示用磷脂酶A2(phospholipase A2)对作为磷脂(phospholipid)的一个种类的磷脂酰乙醇胺(PE)进行处理时生成溶血磷脂酰乙醇胺(lysophosphatidylethanolamine,LPE)的原理的图。
图9是确认用磷脂酶A2对假单胞菌的磷脂提取物进行处理而产生的LPE的结果。用磷脂酶A2对从假单胞菌提取的磷脂进行处理之前(上)和之后(下)利用HPLC-ELSD分析法可以确认PE的分解和LPE的生成。
图10是表示用磷脂酶A2对丁香假单胞菌的磷脂提取物进行处理而产生的LPE主要为LPE16:0和LPE18:1的MS/MS分析结果。
图11中,为了研究在丁香假单胞菌以外的属于假单胞菌属的其它种中是否能够获得成为LPE的原材料的PE,从淤泥假单胞菌(Pseudomonas caeni)、荧光假单胞菌(Pseudomonas fluorescens)、恶臭假单胞菌(Pseudomonas putida)中提取磷脂,结果确认了PE含量在进行实验的假单胞菌的所有种中相当多。
图12是表示用磷脂酶A2对淤泥假单胞菌的磷脂提取物进行处理而产生的LPE主要是LPE18:1和LPE16:0的MS/MS分析结果。
图13是表示用磷脂酶A2对荧光假单胞菌的磷脂提取物进行处理而产生的LPE主要是LPE16:0和LPE18:1的MS/MS分析结果。
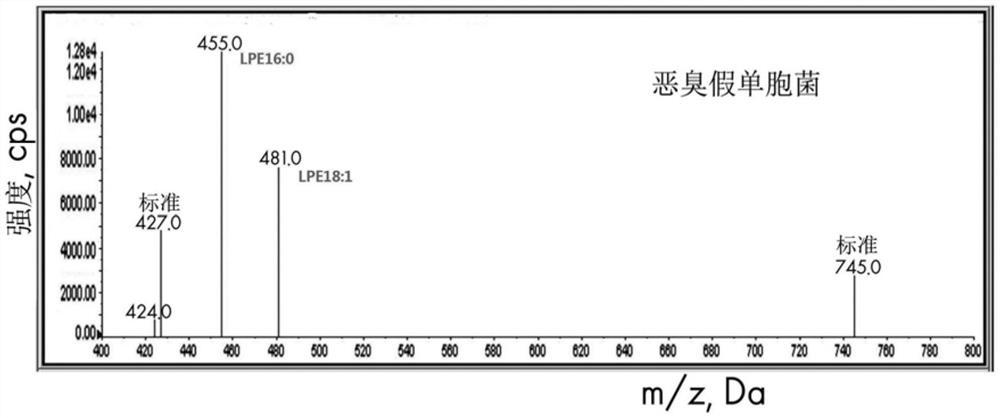
图14是表示用磷脂酶A2对恶臭假单胞菌的磷脂提取物进行处理而产生的LPE主要是LPE16:0和LPE18:1的MS/MS分析结果。
图15中,为了研究在假单胞菌以外的微生物中是否能够获得成为LPE的原材料的PE,从作为革兰氏阳性菌的枯草芽孢杆菌(Bacillus subtilis)、柠檬节杆菌(Arthrobacter citresus)、作为革兰氏阴性菌的大肠杆菌(Escherichia coli)、作为酵母的酿酒酵母(Saccharomyces cerevisiae)、以及作为藻类(Algae)的普通小球藻(Chlorella vulgaris)中提取磷脂。与丁香假单胞菌相比,在大肠杆菌中检出了一定程度的PE含量,确认了在其它微生物中PE含量非常少。
图16是表示用磷脂酶A2对大肠杆菌的磷脂提取物进行处理而产生的LPE主要为LPE16:0且没有LPE18:1的MS/MS分析结果。
具体实施方式
为了实现本发明的目的,本发明提供一种大量生产溶血磷脂酰乙醇胺的方法,其中,包括:
(a)大量培养假单胞菌属(Pseudomonas sp.)微生物后收获所述微生物并从所述微生物提取磷脂的步骤;
(b)用磷脂酶A2处理上述(a)步骤中提取的磷脂以生成溶血磷脂酰乙醇胺的步骤;以及
(c)将上述(b)步骤中生成的溶血磷脂酰乙醇胺进行分离和纯化的步骤。
在根据本发明的一实施方式的方法中,上述溶血磷脂酰乙醇胺(lysophosphatidylethanolamine,以下称为LPE)是利用磷脂酶A2蛋白由侵入植物体的病原菌的细胞膜磷脂产生的,上述LPE可以为LPE 16:0、LPE 16:1、LPE 18:0、LPE 18:1、LPE18:2或LPE 18:3等,可以优选为LPE 16:0、LPE 16:1或LPE 18:1等,可以更优选为LPE 18:1,但不限于此。
另外,在本发明的方法中,上述微生物可以为假单胞菌属(Pseudomonas sp.)微生物,可以优选为淤泥假单胞菌(Pseudomonas caeni)、荧光假单胞菌(Pseudomonasfluorescens)、恶臭假单胞菌(Pseudomonas putida)或丁香假单胞菌(Pseudomonassyringae),可以更优选为淤泥假单胞菌(Pseudomonas caeni)或荧光假单胞菌(Pseudomonas fluorescens),但不限于此。
另外,在本发明的方法中,上述磷脂可以为磷脂酰胆碱(phosphatidyl choline)、磷脂酰丝氨酸(phosphatidylserine)或磷脂酰乙醇胺(phosphati dylethanolamine)等,但可以优选为磷脂酰乙醇胺,可以更优选磷脂酰乙醇胺的1位酰基链为18:1,但并不限定于此。
在根据本发明的一实施方式的方法中,上述磷脂酶A2是将作为生物膜的主要构成成分的甘油磷脂sn-2位的酯键进行水解而使溶血磷脂和脂肪酸游离的酶,本发明的磷脂酶A2可以是来源于蛇毒、蜂毒或猪胰液之类的动物来源、微生物来源或植物来源的酶,另外,也可以是重组的蛋白质,但不限于此。
另外,在本发明的方法中,从获得的细菌中提取磷脂的方法以及分离和纯化溶血磷脂酰乙醇胺的方法可以利用本领域中公知的任意方法。
另外,本发明提供利用上述方法产生的溶血磷脂酰乙醇胺。利用本发明的方法产生的溶血磷脂酰乙醇胺可以优选为溶血磷脂酰乙醇胺18:1,但不限于此。
下面,对本发明的实施例进行详细说明。但是,下述实施例只是用于例示本发明,本发明的内容不限于下述实施例。
材料和方法
植物材料和试剂
拟南芥植物体是在22℃、相对湿度60%中,在16小时光照周期以及光子通量密度(photon flux density)120μmolesm
携带天然启动子::PLA2α的pla2α变异体的互补实验和PLA2α过表达的转基因系统的产生
作为互补(补充),pla2α变异体植物体是通过使用pCAMBIA1300克隆的花序浸渍法(floral dip method)而被转染的,其中所述pCAMBIA1300克隆是包含在BamHI位点具有天然启动子部分(从-1175至+922)的PLA2α基因组DNA的、用于植物转染的二元载体。为了获得用于PLA2α的细胞内易位(不能向质外体(apoplast)分泌)的人工变形,制备了携带着无信号肽的pPLA2α::PLA2α的pCAMBIA1300,并利用土壤杆菌引入到pla2α植物体。为了PLA2α-过表达转基因系统的生成,将具有p35S::PLA2α的pBIG二元载体引入野生型植物体。
植物体的细菌接种
丁香假单胞菌pv.番茄D C 3000(Pseudomonas syringae pv.tomato DC3000)(以下称为Pst)和它的没有毒性的avrRpm1包含菌株Pst-avrRpm1从Y.J.Kim(高丽大学,韩国)获得,没有毒性的avrRpt2包含菌株Pst-avrRpt2从J.M.Park(KRIBB,韩国)获得。没有毒性的avrRps4包含菌株Pst-avrRps4从R.Innes(印第安纳大学,美国)获得。细菌菌株根据Katagiri等(Arabidopsis Book,2002,1:e0039)的方法进行培养并处理。
基因表达分析
总RNA使用RNA分离器(RNA isolator)(Gibco,美国)从冻在液氮中的试样(每个样品为2片叶子)中提取。逆转录聚合酶链反应(RT-PCR)和实时定量聚合酶链反应(real-timeqPCR)使用基因特异性引物而进行。实时定量聚合酶链反应分析使用应用生物系统公司7900 Real-Time PCR System
[表1]
RT-PCR用引物
[表2]
实时qRT-PCR用引物
GUS活性的组织化学性分析和PLA2α向质外体的易位
为了组织化学性位置分析,制备了具有PLA2α-启动子::GUS结构的转基因的拟南芥植物体(Jung et al.,2012,Front Plant Sci.3:126)。为了在叶子组织的感染位置显示出明确的PLA2α表达,在停止光照而转换为夜晚条件的3小时后,将病原菌向PLA2α-启动子::GUS转基因的拟南芥叶的背轴(abaxial)面进行注射器-渗入后,1.5小时后,根据Jefferson(EMBO J.1987,6:3901-3907)等的方法进行了组织化学性GUS分析。
从叶中提取脂质以及ESI-MS/MS分析
总脂质通过以往记载的方法(Ryu et al.,1996,Biochem Biophys Acta.1303:243-250)从喷射了模拟(mock)或Pst-avrRpm1细菌悬浮液(在0.018%Silwet L-77中为1×10
用脂质、水杨酸和重组PLA2α蛋白的处理
所有磷脂从Avanti Polar Lipids Inc.(美国)购买,游离脂肪酸(16:0、18:0和18:1)从Sigma Co.(美国)购买。将溶剂在氮气流(stream)下进行干燥后,通过超声波分解进行悬浮使脂质的最终浓度成为在0.018%silwet L-77中为100nmol ml
重组PLA2α蛋白的成熟的形态在与Dsbc蛋白融合的大肠杆菌中产生,根据以往记载的方法(Ryu et al.,2005,Biochem Biophys Acta.1736:144-151)通过亲和色谱法进行纯化。PLA2α和Dsbc分离的蛋白质混合物或Dsbc:PLA2α融合蛋白(10μgml
PLA2α的抗菌活性和从细菌中释放脂质产物
为了测定PLA2α的抗菌活性,将根据柳等(Biochem Biophys Acta.2005,1736:144-151)的方法产生的3μg游离成熟形态的重组PLA2α添加到悬浮有毒性的Pst(5×10
水杨酸、乙烯和茉莉酸的定量化和NPR1的转换
水杨酸、茉莉酸和乙烯如下定量:根据以往记载的方法(Bowling et al.,1994,Plant Cell 6:1845-1857;Heck et al.,2003,Plant J.36:342-352),由散布Pst-avrRpm1悬浮液(在0.018%Silwet L-77中为1×10
NPR1的转换如下观察:对携带35S::NPR1-eGFP的转基因植物体散布LPE(在0.018%Silwet L-77中为100nmol ml
离子渗出的分析
收获叶组织(每个样品4片叶子),用灭菌水真空渗入5分钟后,一边在常温中搅拌2小时,一边进行培养。样品中的电解质(electrolyte)渗出的程度使用电导率计(conductivity meter,Mettler Toledo,瑞士)测定。显示总电解质渗出的百分比的数据通过冻结后搅拌6小时的同时在常温中进行溶解而获得。
统计
Student's t-test是为了在组间确定统计学意义而使用的。细菌生长分析和离子渗出分析的数据用平均±标准偏差表示,显著性值为*P<0.05、**P<0.01。此外,其它所有数据均用平均±标准误差来表示,*P<0.05、**P<0.01。
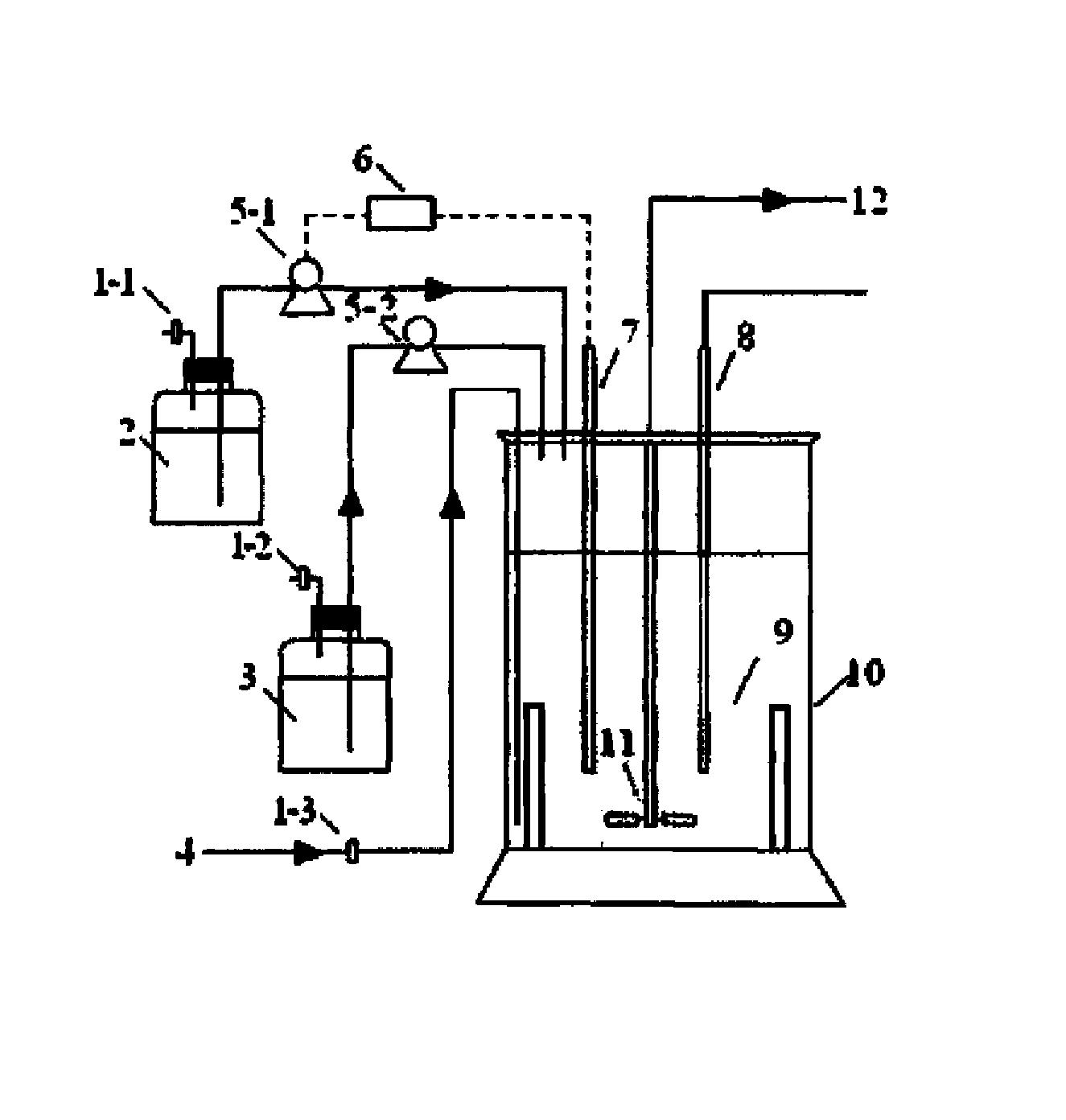
由假单胞菌脂质提取液制备LPE18:1
将假单胞菌细胞培养物进行冻干后,将2mg溶解于水中,进行5秒钟的超声波分解(sonication),得到了假单胞菌基质。此后,将市售的磷脂酶A2(粉末形态)溶于水中,取20μl。将上述假单胞菌基质20μl、磷脂酶A2酶20μl和反应缓冲液(50mM Tris-HCl,10mM CaCl2,0.05%Triton X-100)160μl的混合物在30℃反应30分钟。然后添加750μl的Chl:MeOH(1:2)来停止反应,添加200μl氯仿和200μl的KCl(2M),进行离心分离后,除去上清液,获得了LPE18:1。
实施例1.在pla2α变异体中局部性抗性基因诱导免疫反应的损伤
为了确认在植物防御反应中分泌型PLA2α蛋白的细胞功能,由TAIR获得了作为pla2α变异体的pla2α(敲除)和pla2α-Ⅱ(敲低)。在正常生长条件下,pla2α变异体与野生型植物体相比,在表现型上没有不同。但是,在pla2α变异体中确认了对于Pst-avrRpm1的局部性免疫反应受到了损伤(图1A、1B、1E和1F),并观察到了对于毒性的Pst的pla2α变异体的基本抗性略微受到损伤(图1E)。虽然pla2α变异体对于没有毒性的细菌显示出正常的过敏反应(HR)(图1C),但与野生型植物体相比,在适当地限制细菌生长方面失败了(图1B、1E和1F)。另外,就qRT-PCR分析而言,与野生型相比,pla2α变异植物体显示出显著低的水平的Pst-avrRpm1诱导防御基因表达趋势(图1D)。但是,利用转基因方法对pla2α变异体用天然启动子::pla2α结构补充时,其抑制Pst-avrRpm1生长的能力被恢复(图1E)。pla2α变异体用Pst-avrRpm1接种后,不仅不能开始局部性免疫反应,而且对于Pst-avrRpt2或Pst-avrRps4的反应也不能进行(图1F)。这些结果表明pla2α蛋白与被CC-NB-LRR(卷曲螺旋核苷酸结合位点-富含亮氨酸的重复序列(coiled-coil nucleotide-binding site-leucine-richrepeat))和TIR-NB-LRR(toll-白细胞介素1受体-核苷酸结合-富含亮氨酸的重复序列(toll-interleukin1receptor-nucleotide binding-leucine rich repeat))型抗性基因诱导的免疫反应相关。
实施例2.抗性基因介导的PLA2α的表达和LPE水平的提高
在野生型植物体中,PLA2α以低水平表达。PLA2α表达由于Pst-avrRpm1接种而迅速且适当地被诱导,但是在对于毒性的Pst接种的反应中较弱地被诱导(图2A),从而暗示了PLA2α的表达是由抗性基因介导的。在pPLA2α::GUS分析中也观察到了类似的结果,PLA2α启动子对接种Pst-avrRpm1发生反应而被活化,但是不被毒性的Pst所活化(图2B)。正如所料,与模拟(mock)接种的植物体相比,在接种了Pst-avrRpm1的野生型植物体中,在接种3小时后检测到了诸如LPE的PLA2α的脂质产物的大量增加(图2C)。检测到了LPG(溶血磷脂酰甘油(lysophosphatidylglycerol))和LPC(溶血磷脂酰胆碱(lysophosphatidylcholine))也略微增加。如果接种了毒性的Pst,则不能检测到LPE的增加,检测到只有LPC和LPG略微增加(图2C)。这样的结果表示增加了的LPE的产生是对于没有毒性的Pst的接种的特异性反应。相反地,pla2α变异体在对于Pst-avrRpm1接种的反应中没有显示出增加的LPE的产生(图2D)。与模拟(Mock)处理植物体相比,在被Pst-avrRpm1感染的野生型植物体的叶子中增加的LPE种类为LPE18:1、LPE18:2、LPE18:3和LPE16:1(图2E)。LPE18:1和LPE16:1在模拟(mock)处理植物体几乎没有被检测到,但在Pst-avrRpm1处理叶子中观察到了大量增加(图2E)。有论文报道了LPE18:1和LPE16:1在拟南芥叶组织中是稀有的脂质种类(Devaiah etal.,2006,Phytochemistry 67:1907-1924)。因此,本发明的发明人检测了这种脂质产物是否来源于侵入的细菌的膜。细菌感染刺激PLA2α从细胞质高尔基体向质外体易位。PLA2α显示出对于侵入的细菌的抗菌活性(图2G)。将Pst-avrRpm1悬浮液或Pst-avrRpm1的脂质提取物与重组PLA2α蛋白一同培养时,LPE18:1、LPE16:0和LPE16:1实际上是从细菌或细菌脂质提取物中释放的(图2F)。为了获得关于LPE种类的一部分来源于侵入的病原菌的更直接的证据,将Pst-avrRpm1细菌用[
对野生型叶子接种Pst-avrRpm1时,LPE水平与存在于模拟(mock)处理叶的量相比,增加35%(图2C)。对pla2α变异植物体的叶子接种细菌2.5小时后用重组PLA2α进行渗入的情况下,可以观察到30分钟后在叶子中LPE水平增加100%左右(图2D)。
实施例3.免疫反应中的pla2α变异体的缺陷和外源性LPE18:1处理带来的免疫反应恢复
基于与野生型比较的pla2α变异植物体的分析,我们假设了pla2α变异植物体的免疫反应的缺陷是由介导下游免疫反应的脂质产物生成的失败而引起的。为了确认该假设,Pst-avrRpm1接种1.5小时后,用LPE18:1对pla2α变异体进行处理而补充。实际上,LPE恢复了pla2α变异体诱导局部性免疫反应的能力并限制了疾病症状在所述过敏性反应区域外的传播扩展能力(图3A和3B)。非常重要的是,LPE自身在不存在无毒性的Pst的情况下增加了局部性免疫(图3C-3E)。LPE虽然诱导了作为PR基因的PR1和PDF1.2的表达,但是没有诱导作为伤口相关基因的VSP1和JMT的表达(图3C)。在不存在细菌的情况下LPE导致的局部性免疫的获得通过如下结果而得到证明:与模拟(mock)处理组相比,不仅在野生型植物体中,而且在pla2α变异植物体中,LPE处理抑制了毒性的Pst的细菌生长(图3D)。而且,可以确认与模拟(mock)处理组相比,疾病的症状在散布了LPE的叶子中明显得到缓解(图3E)。这样的结果支持了pla2α变异体的缺陷是由诸如触发下游免疫反应的LPE18:1的脂质介导剂的产生的失败导致的假设。
实施例4.PLA2α与LPE介导的ICS1和NPR1依赖性水杨酸信号传导过程
ICS1位于色素体,在宿主的免疫反应中作为水杨酸生物合成的主要酶而发挥作用。对pla2α变异植物体接种Pst-avrRpm1时,与野生型植物体相比,ICS1的表达减少(图4A)。另外,用Pst-avrRpm1接种的pla2α变异植物体与野生型相比,水杨酸的水平非常低(图4B)。本发明的发明人做出了如下假设:没有生成包含LPE的脂质产物的pla2α变异体在应对没有毒性的细菌攻击的反应中不能表达ICS1,这种缺陷降低水杨酸的水平,导致不能引起导致PR基因表达的下游信号级联放大。为了检验上述假设,确认了外源性的LPE处理是否诱导ICS1表达。用LPE18:1处理的拟南芥叶在处理后第4小时显示出ICS1基因表达,但是不能增加PAL1表达(图4C)。与对照组相比,ICS1基因表达使水杨酸生成增加了2.5倍(图4D)。这样的结果意味着LPE主要通过诱导ICS1-介导的水杨酸生物合成而提高水杨酸水平。
NPR1是导致PR1基因表达的水杨酸介导的免疫反应的重要调节剂。由于水杨酸诱导的氧化还原变化,NPR1活化要求NPR1从细胞质向细胞核易位。由于NPR1活化是由水杨酸介导的,因此LPE诱导的水杨酸的增加可能会使NPR1活化。通过诱导其从细胞质向细胞核的易位(正如水杨酸处理),对携带35S::NPR1-eGFP的转基因植物体应用LPE确实活化了NPR1(图4E)。以上的结果证明了LPE介导了抗性基因诱导的水杨酸生物合成后,伴随着随后的NPR1活化和PR1基因表达的假设。
实施例5.PLA2α与LPE介导的乙烯依赖性防御信号传导过程
pla2α变异体在应对Pst-avrRpm1接种的PDF1.2的表达中也是有缺陷的(图1D),已知其是被茉莉酸和/或乙烯所介导。在对Pst-avrRpm1接种的反应中,pla2α变异植物体中作为乙烯生物合成的ACC合酶(ACS)的基因的表达也受到损伤,并且乙烯的产生减少了(图5A和5B)。与此相反,变异植物体与野生型相比,在茉莉酸的水平上没有大的差异,显示出了LOX2没有被弱化的表达(图5A和5C)。经LPE处理的拟南芥叶显示出ACS基因表达,并显示出乙烯水平的增加,但是在LOX2基因表达或茉莉酸水平上没有确认到明显的增加(图5D-5F)。以上的结果意味着在局部性免疫反应期间,乙烯依赖性的PDF1.2的表达由PLA2α及其脂质产物介导。
实施例6.植物免疫反应中发生的来源于PLA2α的脂质信号传导机制
前面叙述的结果均一致地支持pla2α变异植物体的局部性宿主免疫反应的缺陷是由诸如LPE(其介导PLA2α的抗性基因诱导免疫反应)的脂质产物产生的失败导致的假设。为了支持上述假设,对于pla2α变异体的叶子的重组PLA2α蛋白的外部处理产生了包含LPE的内源性的脂质产物(图2D),诱导了ICS1和PR1基因的原位表达(图6C)。相反地,利用被PLA2抑制剂曼诺力得(Manoalide)催化灭活的重组PLA2α蛋白的处理在诱导PR1基因表达时失败了(图6D)。
以上述结果为基础,可以提出基于PLA2α衍生的脂质信号如下地工作。一旦用无毒性的Pst接种,PLA2α以抗性基因介导的方式快速、适中且暂时性地表达。PLA2α被释放至细胞间空间,在这里其从侵入的细菌和宿主的膜产生包括LPE的脂质产物。然后,通过导致PR1基因表达的ICS1/NPR1-依赖性的水杨酸信号体系和导致PDF1.2基因表达的ACS-依赖性的乙烯信号体系,LPE促进了抗性基因诱导的下游免疫反应(图7)。
实施例7.确认由假单胞菌脂质提取液制备LPE18:1
大量培养丁香假单胞菌(Pseudomonas syringae)并收获后提取脂质。用磷脂酶A2(phospholipase A2)对这种磷脂提取物进行处理后,利用HPLC-ELSD分析法确认了是否PE(磷脂酰乙醇胺(phosphatidylethanolamine))被分解并生成LPE(溶血磷脂酰乙醇胺(lysophosphatidylethanolamine))(图9)。PLA2处理前,只能看到9分钟保留时间(retention time)的PE峰,略微看得到12.5分钟保留时间的LPE峰(图9的上端色谱图),但是PLA2处理后,9分钟保留时间的PE峰消失,出现了12.5分钟保留时间的新的LPE峰,将其用框表示(图9的下端色谱图)。因此,可知用磷脂酶A2对从丁香假单胞菌分离的磷脂进行处理,则生成LPE。通过MS/MS分析,可以确认用PLA2对丁香假单胞菌进行处理后生成的LPE的主要构成成分为LPE18:1和LPE16:0(图10)。
另外,为了了解由其它微生物是否能够产生LPE18:1,从淤泥假单胞菌(Pseudomonas caeni)、荧光假单胞菌(Pseudomonas fluorescens)、恶臭假单胞菌(Pseudomonas putida)中提取磷脂的结果,成为LPE的原材料的PE含量在进行实验的假单胞菌的所有种中均相当多(图11),用PLA2酶进行处理时,确认了大量生成LPE18:1(图12至图14)。
实施例8.检验在假单胞菌以外的其它微生物中的PE产生可能性
为了了解由假单胞菌以外的其它微生物是否也能够产生LPE18:1,从作为革兰氏阴性菌的大肠杆菌(Escherichia coli)、作为革兰氏阳性菌的枯草芽孢杆菌(Bacillussubtilis)、柠檬节杆菌(Arthrobacter citresus)、以及作为酵母的酿酒酵母(Saccharomyces cerevisiae)和作为藻类的普通小球藻(Chlorella vulgaris)中分离磷脂并进行分析的结果,除了大肠杆菌以外的其它微生物中,几乎检测不到作为LPE的原材料的PE(图15)。虽然在大肠杆菌中提取了一定程度的PE,但是用PLA2进行处理的结果,只生成了LPE16:0(图16)。因此,可知在假单胞菌以外的其它微生物中,至少在研究过的微生物中,难以确保能够生成LPE18:1的PE。
<110> (韩国生命工学研究院)
<120> 从假单胞菌属微生物生产溶血磷脂酰乙醇胺18:1的方法
<130> PCT1511
<160> 46
<170> KopatentIn 2.0
<210> 1
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 1
ggcgatgaag ctcaatccaa acg
<210> 2
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 2
ggtcacgacc agcaagatca agac
<210> 3
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 3
ggggataagg ggttctcaca
<210> 4
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 4
ctgccctagt tacaacccga
<210> 5
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 5
ggccaaagag ggtatcatcg
<210> 6
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 6
gctcgaccac agctcttatg g
<210> 7
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 7
aaagaacatg gtgatcaacg c
<210> 8
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 8
agttgagatc gcagccactt
<210> 9
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 9
cacccttatc ttcgctgctc 20
<210> 10
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 10
gttgcatgat ccatgtttgg
<210> 11
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 11
cttaacgtcg gtgttcagct c 21
<210> 12
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 12
gggtttcttg aggactttgc c 21
<210> 13
<211> 18
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 13
tcgcacttca ttgatgcg 18
<210> 14
<211> 28
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 14
tcatagctct gttttcatat cattacct 28
<210> 15
<211> 18
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 15
gtcacgtgtt gctttcgg 18
<210> 16
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 16
aacgtttgaa ctgcttgtg 19
<210> 17
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 17
gctttaggct taaccgtctt 20
<210> 18
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 18
agaaggagaa gggttcatc 19
<210> 19
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 19
gtgctcttgt tcttccctcg 20
<210> 20
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 20
aaggcccacc agagtgtatg 20
<210> 21
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 21
ctcatactca agccaaacgg atc 23
<210> 22
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 22
gccatgaaga tagatgctta att 23
<210> 23
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 23
acctcttctc cgagcatgaa 20
<210> 24
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物
<400> 24
gccgtcaaaa acaaccctaa 20
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。


















动态评分
0.0