








专利摘要
本发明的特征在于作为疫苗靶标且作为对抗播散性念珠菌病的预防策略的HYR1。
说明书
关于联邦资金资助研究的声明
本发明是在政府的支持下由公共卫生基金(Public Health Service grants)R21AI066010、R01 AI067703、R01 AI063503和R03 AI083251资助完成的。美国政府对本发明享有一定的权利。
技术领域
本发明涉及念珠菌(Candida)菌丝细胞壁蛋白,由对用念珠菌菌丝细胞壁蛋白免疫接种的免疫应答产生的抗体,以及预防和/或治疗念珠菌病和念珠菌菌丝细胞壁蛋白引起的其他细菌感染的方法。
背景技术
本文提及的所有文件,或指示的部分通过引用并入本文,包括于2009年7月3日提交的美国临时申请号61/223,005。然而,并没有承认这些文件是本发明要求保护的主题的现有技术。
在美国每年发生大约60,000例播散性念珠菌病[1],导致数百亿美元的卫生保健费用。考虑到这种感染有40%的死亡率,需要鉴定新的用于干预的预防或治疗靶标。
对抗播散性念珠菌病的主要宿主防御机制是生物体的吞噬杀伤作用[2,3]。仅吞噬细胞能够在体外直接杀死念珠菌[4]。另外,在小鼠、兔、犬或人中静脉接种念珠菌的30分钟内,酵母菌仍然留在网状内皮系统内,尤其是在肝内。富含枯否巨噬细胞(Kupffer macrophage)的肝脏在单程期间能够清除门脉系统中99.9%的酵母菌[5],强调了吞噬防御机制对抗真菌的有效性。因此,白色念珠菌(C.albicans)对吞噬杀伤作用的抗性是该生物体的重要致病功能。
细胞表面糖基化磷脂酰肌醇(GPI)-锚定蛋白处于病原体和宿主之间的关键界面处,使得这些蛋白质可能参与宿主-病原体相互作用[6]。
该生物体的产生毒力的调控途径中的效应器的鉴定为使用优于现有抗真菌剂的方法或组合物进行治疗干预提供了机会。影响涉及毒力的调控途径的细胞表面蛋白或菌丝蛋白的鉴定是特别有希望的,因为所述蛋白质的表征能够获得可能优于现有抗真菌剂或与现有抗真菌剂协同的对抗念珠菌感染的免疫治疗技术。
白色念珠菌的毒力受几种推定的毒力因子调控,所述毒力因子与宿主成分的粘附及其从酵母-至-菌丝的转化能力在决定致病性方面是最关键的。尽管存在有杀灭念珠菌的有效抗真菌剂,但由念珠菌血症导致的死亡率仍然为大约38%,甚至采用有效的抗真菌剂诸如两性霉素B治疗时也是如此。此外,现有的药剂诸如两性霉素B往往显示不合乎需要的毒性。尽管可以开发出比两性霉素B毒性更小的其他抗真菌剂,但不可能开发出更有效的药剂。因此,治疗或预防播散性念珠菌病的被动或主动免疫治疗是标准抗真菌治疗的有前途的替代方法。
因此,存在着对有效免疫原的需求,所述免疫原将提供针对念珠菌和其他免疫原性相关的病原体的宿主免疫保护和被动免疫保护。本发明满足了此需求并且还提供了相关的优势。
发明内容
本发明的特征在于念珠菌HYR1多肽抗原,和所述抗原的治疗用途。本发明的HYR1多肽抗原可以用来治疗或预防受试者中的念珠菌感染。
通过以白色念珠菌(C.albicans)中的GPI-锚定蛋白为中心筛选了新的条件性过表达/抑制系统,我们鉴定了作为毒力因子的HYR1。HYR1是菌丝共表达基因,其缺失突变株在体外不显示任何形态异常[7]。下面我们提供了结果证明HYR1在体外介导对吞噬杀伤的抗性,在体内调节组织真菌载量,并且因此是改善播散性念珠菌病的严重性的疫苗靶标。
定义
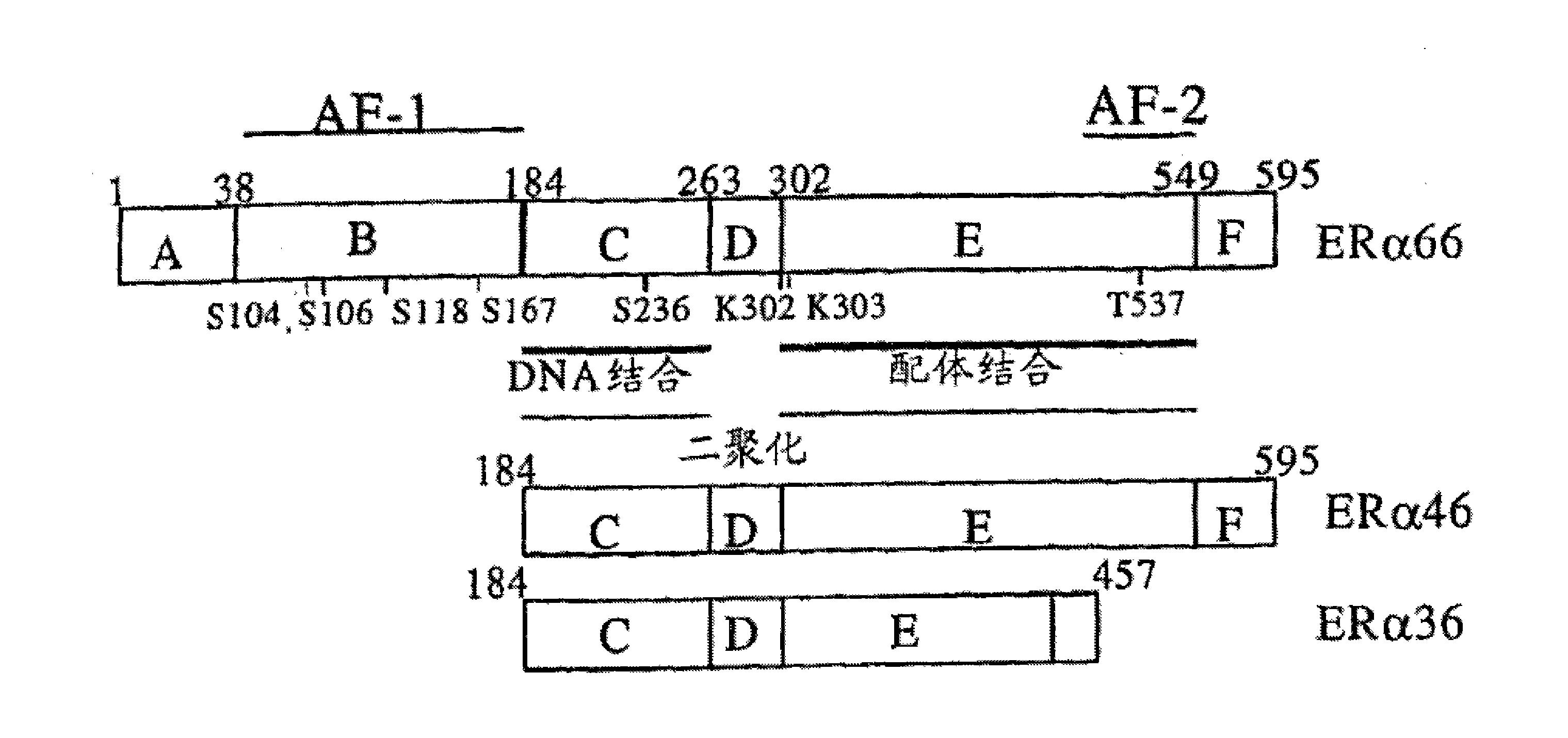
“HYR1”多肽是指与SEQ ID NO:1的氨基酸序列基本相同的多肽。期望地,HYR1多肽与SEQ ID NO:1的氨基酸序列具有至少70、75%、80%、85%、90%、95%、99%或甚至100%的同一性。
“HYR1多肽片段”或“HYR1片段”是指HYR1多肽的片段,该片段包含少于937、936或935个氨基酸。优选的HYR1片段的长度为300至350或250至500个氨基酸。期望地,所述片段少于937、936、935、934、933、932、931或930、920、910、900、890、880、870、860、850、840、830、820、810、800、790、780、770、760、750、740、730、720、710、700、690、680、670、660、650、640、630、620、610、600、590、580、570、560、550、540、530、520、510、500、490、480、470、460、450、440、430、420、410、400、390、380、370、360、350、340、330、320、310、300、290、280、270、260、250、240、230、220、210、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、25、20、15或10个氨基酸,并且期望是免疫原性的。例如,HYR1片段可以包含SEQ ID NO:2序列中的一个或多个保守氨基酸置换。其他期望的HYR1片段包含SEQ ID NO:2序列中的一个或多个保守氨基酸置换和/或SEQ ID NO:2序列的N-和/或C-末端处的至少一个侧翼氨基酸(例如,1、2、3、4、5、6、7、8、9或10个侧翼氨基酸)。其他优选的HYR1片段包含SEQ ID NO:2序列的7个以上的连续氨基酸。
HYR1片段的非限制性实例包括SEQ ID NO:2序列的氨基酸1-40、10-50、20-60、30-70、40-80、50-90、60-100、70-110、80-120、90-130、100-140、110-150、120-160、130-170、140-180、150-190、160-200、170-210、180-220、190-230、200-240、210-250、220-260、230-270、240-280、250-290以及260-300、270-310、280-320和290-331氨基酸;和具有下列特征中的一个或多个的这些片段:SEQ ID NO:2序列中的一个或多个保守氨基酸置换(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、或16个保守氨基酸置换);从SEQ ID NO:2序列的N和/或C-末端截去一个或多个氨基酸(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15或16个氨基酸);和SEQ ID NO:2序列的N-和/或C-末端处的至少一个侧翼氨基酸(例如,1、2、3、4、5、6、7、8、9或10个侧翼氨基酸)。
“基本相同”是指显示与参照氨基酸序列具有至少50%、期望60%、70%、75%或80%、更期望85%、90%或95%、和最期望99%氨基酸序列同一性的多肽。比较序列的长度通常为至少10个氨基酸、期望至少15个连续氨基酸、更期望至少20、25、50、75、90、100、150、200、250、275、300、310、315、320、325、330、335、340、345或350个连续氨基酸,和最期望全长氨基酸序列。
序列同一性可以使用缺省设置的序列分析软件(例如,Genetics Computer Group的序列分析软件包,威斯康星大学生物技术中心(University of Wisconsin Biotechnology Center),1710 University Avenue,Madison,WI 53705)来测量。此类软件可以通过指定与各种置换、缺失和其他修饰的同源性程度来匹配相似序列。还可以使用Clustal W(1.4)程序(由德国欧洲分子生物学实验室(European Molecular Biology Laboratory,Germany)的Julie D.Thompson和Toby Gibson以及英国剑桥欧洲生物信息学协会(European Bioinformatics Institute,Cambridge,UK)的Desmond Higgins编制)通过设置配对比对模式为“slow”,设置配对比对参数以包括10.0的开放空位罚分和0.1的延伸空位罚分,以及设置相似性矩阵为“blosum”,延迟比对(delay divergent)为40%,和空位距离为8,来比对多条序列。
“保守氨基酸置换”当在本文中使用时是指在氨基酸序列中的一个氨基酸替换为其侧链的化学性质相关的氨基酸家族内的另一个氨基酸。
遗传编码的氨基酸可以分成4个家族:酸性氨基酸(天冬氨酸、谷氨酸);碱性(赖氨酸、精氨酸、组氨酸);非极性氨基酸(丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸);和不带电荷的极性氨基酸(甘氨酸、天冬酰胺、谷氨酰胺、半胱氨酸、丝氨酸、苏氨酸、酪氨酸)。苯丙氨酸、色氨酸和酪氨酸有时归类为芳族氨基酸。以相似的方式,氨基酸也可以分成以下各组:酸性氨基酸(天冬氨酸、谷氨酸);碱性氨基酸(赖氨酸、精氨酸、组氨酸);脂族氨基酸(甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、丝氨酸、苏氨酸),丝氨酸和苏氨酸任选地被单独归类为脂族-羟基氨基酸;芳族氨基酸(苯丙氨酸、酪氨酸、色氨酸);酰胺氨基酸(天冬酰胺、谷氨酰胺);和含硫氨基酸(半胱氨酸、甲硫氨酸)。
氨基酸序列的改变是否产生功能同源物可以使用标准方法诸如本文所述的测定通过评估变体肽以类似于野生型蛋白相似的方式起作用的能力来确定。
本发明期望的实施方案包括SEQ ID NO:1或2的氨基酸序列中的至少一个保守氨基酸置换;和更期望SEQ ID NO:1或2的序列中的1、2、3、4、5、6、7、8、9或10个保守氨基酸置换。
“侧翼氨基酸”是指多肽序列中与特定序列的N-或C-末端紧邻的氨基酸。期望地,侧翼氨基酸存在于SEQ ID NO:1或2或其片段的氨基酸序列的N-和/或C-末端上;和更期望地,1、2、3、4、5、6、7、8、9或10个侧翼氨基酸存在于SEQ ID NO:1或2或其片段的氨基酸序列的N-和/或C-末端处。
当在本文中使用时,“融合蛋白”是指由(1)HYR1多肽、HYR1片段;和(2)融合伴侣组成的多肽。
当在本文中使用时,“融合伴侣”是指可以与HYR1多肽或HYR1片段融合的异源序列。融合伴侣的实例在本文中描述并且包括检测标志物、稳定结构域或有助于蛋白质的产生或纯化的序列。
当在本文中使用时,“免疫应答”是指生物体的免疫系统响应于抗原或传染原的活化。在脊椎动物中,这可以包括,但不限于,下列中的一种或多种:幼稚B细胞成熟为记忆B细胞;浆细胞(效应B细胞)产生抗体;诱发细胞介导免疫;CD4+T细胞引起的活化和细胞因子释放;CD8+T细胞活化和引起的细胞因子释放;细胞因子募集和吞噬细胞(例如,巨噬细胞、中性粒细胞、嗜酸性粒细胞)活化;和/或补体活化。
“免疫原性的”是指能够在受试者中诱发免疫应答的任何物质。
“药学可接受的盐”是指在制药工业中使用的任何无毒的酸加成盐或金属络合物。酸加成盐的实例包括有机酸诸如乙酸、乳酸、扑酸、马来酸、柠檬酸、苹果酸、抗坏血酸、琥珀酸、苯甲酸、棕榈酸、辛二酸、水杨酸、酒石酸、甲磺酸、甲苯磺酸或三氟乙酸等;高分子酸诸如丹宁酸、羧甲基纤维素等;和无机酸诸如盐酸、氢溴酸、硫酸、磷酸等。金属络合物包括锌、铁等。
“药学可接受的载体”是指用来溶解和递送药剂至受试者的任何溶液。合乎需要的药学可接受的载体是盐水。在期望的实施方案中,药学可接受的载体包括佐剂。示例性的佐剂在本文中描述。其他生理学可接受的载体和它们的制剂是本领域技术人员已知的,并且描述在,例如,Remington’s Pharmaceutical Sciences,(第20版),A.Gennaro编辑,2003,Lippincott Williams & Wilkins中。
“分离的”是指已经与其天然伴随的成分分离的蛋白质(或其片段)。典型地,当多肽的至少60重量%不含有与其天然结合的蛋白质和天然存在的有机分子时,则该多肽是基本分离的。该定义还延伸至与其侧翼氨基酸分离的多肽(例如,对于氨基酸序列,分离的是指不含有侧翼氨基酸的序列,在多肽中所述侧翼氨基酸与所述序列天然地结合)。优选地,所述多肽的至少75重量%、更优选至少90重量%和最优选至少99重量%被分离。分离的多肽可通过标准技术获得,例如,通过从天然来源提取(例如,从用念珠菌感染的细胞中纯化),通过编码HYR1片段或其融合蛋白的重组核酸的表达;通过化学合成所述多肽。纯度可以通过任何适宜的方法测量,例如,通过柱色谱、聚丙烯酰胺凝胶电泳,或HPLC分析。
“治疗有效量”是指在受试者中产生下列的一种或多种效应所需的免疫原性化合物(例如,多肽、片段、融合蛋白或疫苗)的量:免疫应答;念珠菌感染水平的下降(例如,减少至少5%、10%、20%或30%;更期望40%、50%、60%或70%;和最期望80%或90%);患者中念珠菌感染的一种或多种症状的减轻(例如,减轻至少10%、20%、30%、40%、50%、60%、70%、80%、90%或甚至100%);或对新念珠菌感染的抵抗力增加(例如,增加至少5%、10%、20%、30%、40%或50%;更期望60%、70%、80%、或90%;或最期望100%、200%或300%)。
从下面的详述、附图和权利要求,本发明的其他特征和优势将是显而易见的。
附图说明
图1.HYR1的条件性表达增强中性粒细胞和巨噬细胞介导的对白色念珠菌的杀伤。(A)HYR1条件性过表达/抑制株CAAH-31的证实。HYR1的RT-PCR结果证明该基因在-DOX培养基中的过表达和在+DOX培养基中缺少表达。EFB1片段被共同扩增并作为对照。缺少包含内含子EFB1的919bp条带证实cDNA制剂中没有基因组DNA污染。THE31是野生型对照菌株。(B)白色念珠菌在含有DOX(抑制HYR1)和不含DOX(过表达HYR1)的YPD中在30℃下生长过夜,然后与人中性粒细胞共培养;(C)与HL-60来源的中性粒细胞共培养的白色念珠菌;(D)与HL-60来源的巨噬细胞共培养的白色念珠菌。
图2.白色念珠菌HYR1的表达增加了bcr1缺失突变体或光滑念珠菌(C.glabrata)菌株中对人中性粒细胞杀伤的抵抗力。(A和B)HYR1在白色念珠菌的bcr1缺陷株中的自主表达完全补偿了亲本株对中性粒细胞杀伤抵抗力的超敏性。白色念珠菌菌株DAY185(野生型)、CJN702(bcr1缺失突变株)和CJN698(BCR1补偿株)CJN114、CJN1153、CJN1222、CJN1259、CJN1276、CJN1281和CJN1288(在bcr1缺失突变体背景中分别自主表达ALS1、ALS3、HWP1、HYR1、RBT5、CHT2和ECE1)在YPD中在30℃下生长过夜。数据显示为中值±四分位数。*P<0.04相对于野生型和BCR1-补偿株。(C)白色念珠菌HYR1基因的异源表达增加了光滑念珠菌对HL-60来源的中性粒细胞介导的杀伤的抵抗力。*P<0.0001.
图3.检测播散性念珠菌病期间的HYR1表达,尽管在体外其表达最初被中性粒细胞抑制。(A)在用白色念珠菌静脉内感染后6或24h收获肾、肝、肺、脾和脑。巢式RT-PCR用来检测HYR1的表达。白色念珠菌EFB1和小鼠看家基因G3PDH用作对照。+表示感染的小鼠,而-表示未感染的小鼠。(B)HL-60来源的中性粒细胞抑制白色念珠菌HYR1表达。野生型白色念珠菌在添加了10%混合人血清的RPMI 1640中培养。在不添加中性粒细胞的情况下,在半小时诱导后检测到HYR1表达。添加中性粒细胞,抑制HYR1表达两小时。
图4.HYR1的体内表达及其对真菌载量的作用。为了在播散性念珠菌病期间检测HYR1表达,在用白色念珠菌静脉内感染后6或24h收获肾、肝、肺、脾和脑。巢式RT-PCR用来检测HYR1的表达。白色念珠菌EFB1和小鼠看家基因G3PDH用作对照。+表示感染的小鼠,而-表示未感染的小鼠(A)。HYR1的条件性表达增加了具有大量组织吞噬细胞的器官中的真菌载量。在用白色念珠菌HYR1或生长在条件性表达(-DOX)或抑制性(+DOX)条件下的对照菌株感染的免疫感受态小鼠(n=8只/组)的肝和脾中的白色念珠菌载量(A)和体内HYR1表达(B)。感染后1天收获肝和脾。y-轴反映了测定的检测下限。数据显示为中值±四分位数。*P<0.0001相对于HYR1株(HYR1-DOX)或对照菌株的不表达。
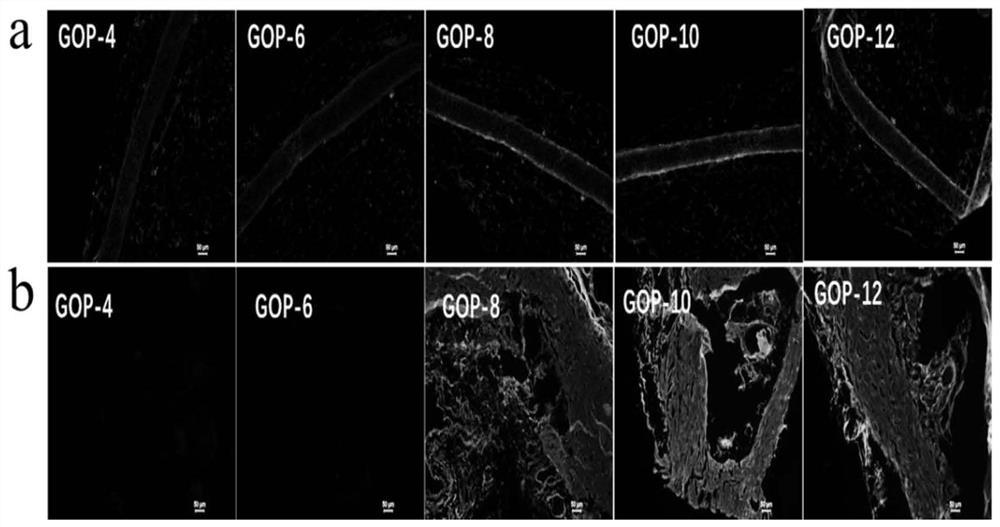
图5.采用抗-Hyr1p血清的间接免疫荧光证明了白色念珠菌菌丝上Hyr1p的表面表达。通过将白色念珠菌在RPMI 1640中孵育90min诱导菌丝形成。细胞用hyr1缺失突变体预吸收的抗-Hyr1p血清(1∶100)或来自空载质粒克隆血清的抗蛋白制剂(阴性对照)染色,然后用Alexa标记的抗小鼠Ab染色。
图6.Hyr1p(rHyr1p-N)的重组N-末端明显地防止鼠血原性播散性念珠菌病。(A)用混合有完全或不完全弗氏佐剂的rHyr1p-N免疫接种并通过尾静脉用2.2x105个白色念珠菌SC5314的芽生孢子感染的小鼠(8只小鼠/组)的存活率。(B)用混合有0.1%的铝胶(alhydrogel)的rHyr1p-N或脱毒的rHyr1p-N免疫接种并用7x105个白色念珠菌15563的芽生孢子感染的小鼠(8只小鼠/组,例外是对照组有17只小鼠)的存活率。通过对数秩检验(log-rank test)*P=.001。(C)免疫接种的或对照F(ab)’2对白色念珠菌条件性表达或抑制的Hyr1阻断小鼠中性粒细胞杀伤的效应。对照表示在缺少任一种F(ab)’2的条件下进行的测定。数据显示为中值±四分位距。通过Mann-Whitney检验*P=.001。
具体实施方式
白色念珠菌是人类的一种常见病原体。例如,白色念珠菌(C.albicans)尽管一般是一种无害的共生菌,也可以导致多种病症,从浅表皮肤粘膜感染诸如阴道和/或口咽念珠菌病至播散性念珠菌病中累及的深部器官。在导致疾病前,真菌定植在胃肠道,并且在一些情况下定植在皮肤和粘膜中。与宿主粘膜表面的粘附是此初始步骤的关键必要条件。在定植后,白色念珠菌经由感染的静脉内装置或通过被化疗或应激性溃疡破坏的胃肠粘膜的移行而进入血流。生物体经血流播散,结合并穿透血管内皮从而从血管树中移出,并侵入深部器官诸如肝、脾和肾。
本文所述的HYR1片段的鉴定和功能表征使得此多肽将被有效地用于念珠菌病的治疗中。
白色念珠菌通过与内皮细胞的粘附的发病机制性质在美国专利5,578,309中进行了讨论,该专利的全部内容具体地通过引用并入本文。为了描述HYR1基因及其特征,包括基因产物的表征,参见,Bailey等人,(Journal of Bacteriology178:5353-5360,1996)。
本发明提供了一种疫苗,其具有处于药学可接受的介质中的分离的HYR1片段和任选的佐剂。所述疫苗可以是来源于念珠菌物种的HYR1片段,所述念珠菌物种诸如白色念珠菌(Candida albicans)、克鲁斯念珠菌(Candida krusei)、热带念珠菌(Candida tropicalis)、光滑念珠菌(Candida glabrata)、或近光滑念珠菌(Candida parasilosis)。
本发明利用白色念珠菌HYR1序列的基因产物作为治疗、预防或减轻播散性念珠菌病的疫苗。该疫苗有效地对抗白色念珠菌的不同菌株以及对抗不同的念珠菌属。
因此,根据一个方面,本发明提供了一种HYR1片段,当其配制成药学组合物并作为含有或不含佐剂的疫苗施用时是有用的。HYR1片段可以是念珠菌来源的并且可以例如,由属于念珠菌属的物种获得,所述物种例如近光滑念珠菌、克鲁斯念珠菌、光滑念珠菌和热带念珠菌。HYR1片段可以以分离或纯化的形式获得,并且因此,根据本发明的一个实施方案,HYR1片段配制为疫苗以导致患者中的免疫应答以引发针对念珠菌的免疫应答。
本发明还提供了一种治疗或预防播散性念珠菌病的方法。该方法包括施用致免疫量的HYR1片段的疫苗。疫苗可以与佐剂或不与佐剂一起施用。HYR1片段可以来源于不同的念珠菌菌株以及来自不同的念珠菌物种诸如白色念珠菌、克鲁斯念珠菌、热带念珠菌、光滑念珠菌和近光滑念珠菌。
本发明的疫苗针对不同的念珠菌菌株、不同的念珠菌物种、其他细菌和传染原的有效性和它们广泛的免疫活性根据标准方法诸如下面进一步描述的那些来评估。
考虑到本文提供的教导和指导,本领域技术人员将理解本领域公知的免疫治疗方法可以采用处于药学可接受的组合物中的一种或多种HYR1片段,所述组合物作为含有或不含佐剂的疫苗施用。为了本发明的目的,术语“药学的”或“药学可接受的”是指通过已知技术配制的组合物,该组合物将是无毒的,并且当需要时,与可以安全地施用于人类的载体或添加剂一起使用。可以使用公知的途径进行施用,所述途径包括,例如,静脉内、肌肉内、腹膜内或皮下注射。本发明的这种疫苗还可以包含本领域技术人员已知的缓冲剂、盐或其他溶剂以保持溶液中疫苗的活性。同样,本领域中公知的大量佐剂中的任一种可以与本发明的疫苗一起使用以诱发、促进或增强治疗有效的免疫应答,所述免疫应答能够减少或阻断念珠菌与易感宿主细胞的结合、对易感宿主细胞的侵袭和/或感染。
要理解的是基本不影响本发明的各种实施方案的活性的改动也包括在对本文提供的发明的定义之内。因此,下面的实施例意在说明而非限制本发明。
实施例I
白色念珠菌Hyr1p赋予对中性粒细胞杀伤的抵抗力并且是疫苗靶标
如上所讨论,白色念珠菌是人类侵袭性真菌感染的最常见病因。尚不清楚白色念珠菌在威胁生命的全身感染期间在恶劣的血液环境中如何逃脱吞噬细胞的攻击和存活。使用条件性过表达/抑制遗传策略,我们发现HYR1基因在体外减少吞噬细胞对白色念珠菌的杀伤并且在体内增加组织真菌载量。与转录因子Bcr1p的正性调节一致,在体外HYR1补偿了白色念珠菌的bcr1缺失突变体对吞噬细胞介导的杀伤的超敏性。此外,光滑念珠菌中HYR1的异源表达使得生物体对中性粒细胞杀伤具有更大的抵抗力。最后,使用重组Hyr1p免疫接种显著地保护小鼠免受血原性播散性念珠菌病。因此,Hyr1是白色念珠菌的重要毒力因子,其介导对吞噬细胞杀伤的抵抗力。Hyr1p因此是用于改善播散性念珠菌病的结局的疫苗或其他免疫学或小分子干预的靶标。
结果
在体外HYR1在芽生孢子中的条件性表达显著地增强了白色念珠菌对吞噬细胞介导的杀伤的抵抗力
为了研究HYR1的功能,我们构建了白色念珠菌的条件性过表达/抑制菌株,CAAH-31。在CAAH-31中,HYR1的一个等位基因受四环素调节(TR)的启动子控制,且另一个等位基因被破坏。通过半定量RT-PCR,我们证实当CAAH-31的芽生孢子在不含DOX的培养基中生长时HYR1被大量地表达,并且在DOX的存在下则检测不到(图1A)。如期望的那样,在野生型(THE31)芽生孢子中检测不到HYR1,因为HYR1是菌丝共表达基因(图1A)。
HYR1条件性过表达株CAAH-31和THE31野生型对照具有相同的生长速率,无论是否存在DOX(不存在DOX时野生型对照菌株的倍增时间=1.51±0.29h,存在DOX时=1.51±0.38h;不存在DOX时CAAH-31菌株的倍增时间=1.39±0.30h,存在DOX时=1.35±0.19h)。我们还评价了HYR1过表达对细胞表面上其他GPI-锚定蛋白的正常积累的影响。通过直接免疫荧光,我们证实HYR1过表达对GPI-锚定蛋白Als1p的积累没有影响(数据未示出)[8]。
在毒力相关表型的常规筛选期间,我们测定了HYR1的条件性过表达对人吞噬细胞的念珠菌杀伤的影响。表达HYR1的白色念珠菌(CAAH-31-DOX)对人中性粒细胞介导的杀伤的抵抗力明显地高于野生型白色念珠菌(在芽生孢子期不表达HYR1)和HYR1-抑制的白色念珠菌(CAAH-31+DOX)(图1B)。此表型不是由于DOX引起的,因为在野生型对照和HYR1-抑制的白色念珠菌(+DOX)之间没有显著的差异。
我们还使用HL-60细胞系进行了念珠菌杀伤测定,该HL-60细胞系可以分化成中性粒细胞样或巨噬细胞样细胞[9,10]。类似于新鲜收获的人中性粒细胞,HYR1的条件性过表达减少了HL-60中性粒细胞样(图1C)和巨噬细胞样(图1D)细胞两者在体外对白色念珠菌芽生孢子的杀伤。
体外HYR1表达补偿了bcr1缺失突变体白色念珠菌对中性粒细胞杀伤的超敏性
由于HYR1是正性转录调节因子Bcr1p的下游基因[17],我们假设bcr1的破坏将在促进野生型白色念珠菌表达HYR1的条件下加剧对中性粒细胞杀伤的易感性(即,在菌丝形成期间)。因此我们通过将细胞在添加了10%FBS的RPMI中在37℃下孵育40min诱导白色念珠菌形成芽管。此条件已知诱导HYR1的表达[18],并且产生的芽管足够短使得不会形成大量的菌丝,由此在我们的杀伤测定中能够定量菌落形成单位(CFU)。我们比较了bcr1缺失突变株(CJN702)、bcr1缺失突变体背景中的BCR1补偿菌株(CJN698)和野生型白色念珠菌菌株(DAY185)的中性粒细胞杀伤。与BCR1-补偿的和野生型对照菌株相比,bcr1缺失突变体对中性粒细胞介导的杀伤具有超敏性(图2A)。此外,通过bcr1突变体背景中的HYR1的自主表达完全补偿了bcr1-缺陷性白色念珠菌对杀伤的超敏性,而非由Bcr1p调节的其他细胞表面编码基因补偿[17](图2A和2B)。
过度表达HYR1的白色念珠菌对吞噬细胞杀伤的抵抗力可以通过HYR1在光滑念珠菌的异源表达而重现
为了进一步确定由HYR1介导的毒力表型,我们使用携带组成型PGK1启动子的质粒pGRB2.2[11]在光滑念珠菌BG14[19]中异源表达了该基因。我们还生成了用空载质粒转化的光滑念珠菌BG14作为阴性对照。与用空载质粒转化的光滑念珠菌相比,HYR1在光滑念珠菌中的表达导致在体外由HL-60来源的中性粒细胞引起的杀伤减少75%(图2C)。
中性粒细胞抑制念珠菌HYR1表达
由于HYR1的条件性过表达赋予念珠菌以对中性粒细胞介导的杀伤的抵抗力,我们使用RT-PCR来研究在体外野生型白色念珠菌(SC5314)HYR1响应于HL-60-来源的中性粒细胞的表达。早在暴露于包含血清的培养基后30min就表达HYR1,并且在培养期间维持高表达持续2.5h(图3A)。然而,当在培养中白色念珠菌暴露于HL-60-来源的中性粒细胞时,甚至在血清的存在下,就可抑制共培养物中的HYR1表达直至2h(图3A)。
野生型白色念珠菌在血原性播散性念珠菌病期间表达HYR1,导致富含呑噬细胞的器官中的组织真菌载量增加
为了确定在血原性播散性念珠菌病期间HYR1是否表达,在感染6小时和24小时后从用白色念珠菌野生型菌株感染的小鼠中收获5种主要器官-脑、肝、肺、脾和肾。使用改良的巢式-RT-PCR测定[20]来评估体内HYR1表达。在所有5种器官中均检测到HYR1表达(图3B)。
HYR1的过表达增加肝和脾中的真菌载量,而HYR1的抑制减少肝和脾中的真菌载量(图4A)。此外,我们证实与野生型菌株相比,过表达菌株在肝中具有显著较高的HYR1水平;抑制的菌株证明有减小的水平的趋势(图4B)。相反,HYR1表达不显著地改变肾中的真菌载量(数据未示出),肾是缺少常驻吞噬细胞的器官。
作为疫苗候选物的rHyr1p-N
基于序列分析,预测Hyr1p是一种细胞表面蛋白[7]。为了证实此预测,我们在用表达克隆转化的大肠杆菌(E.coli)中生成了重组N-末端Hyr1p,该表达克隆包含编码序列的氨基酸25-350(rHyr1p-N)。来自用rHyr1p-N免疫的小鼠的血清针对白色念珠菌的hyr1缺失突变体进行预吸附[7],然后在野生型菌丝上进行间接免疫染色。我们发现野生型白色念珠菌菌丝的细胞壁强染色(图5),证实其是细胞表面表达的,并且因此暴露于免疫系统。
由于Hyr1p是细胞表面蛋白,其赋予对吞噬细胞的念珠菌杀伤作用的抵抗力,我们力图确定其作为疫苗侯选物的潜力。小鼠用rHyr1p-N结合佐剂或单独佐剂免疫接种。强化免疫后2周,小鼠经尾静脉用高毒力白色念珠菌SC5314感染。与用单独佐剂免疫接种的小鼠相比,用rHyr1p-N免疫接种明显地改善了小鼠的存活率(在35天时存活率分别为62.5%和0%)(图6A)。
与用任一种佐剂单独免疫接种的小鼠相比,用混合有完全或不完全弗氏佐剂或明矾的rHyr1p-N免疫接种明显改善了小鼠的存活率(图6A和6B)。
抗-rHyr1p血清通过直接抑制Hyr1p抗中性粒细胞功能增强了小鼠中的中性粒细胞杀伤。
rHyr1p-N疫苗的保护作用表明,抗-rHyr1p血清可能能够中和Hyr1p在白色念珠菌中的保护作用。为了确定抗-rHyr1p抗体是否能够直接抑制Hyr1p功能,我们从用rHyr1p-N或对照(从用空载质粒转化的大肠杆菌细胞中制备的制剂)免疫接种的小鼠的总IgG中分离和制备了F(ab)’2片段。我们发现来自免疫血清而非对照血清的F(ab)’2能够将HYR1条件性表达菌株的中性粒细胞杀伤恢复至与抑制性菌株相同的水平(图6C)。
总结
在此研究中,我们证明了HYR1,一种菌丝共表达基因[7,21],编码念珠菌吞噬细胞抗性因子。与野生型白色念珠菌芽生孢子相比,HYR1在念珠菌芽生孢子中的条件性表达导致真菌对吞噬细胞的杀伤具有更大的抵抗力。另外地,HYR1在白色念珠菌中的功能通过在体外在光滑念珠菌中异源表达该基因而得到重现。我们还发现缺少Bcr1p(一种正性调节HYR1表达的转录因子[17])的菌株显示增强的对吞噬细胞介导的杀伤的易感性。bcr1缺失突变体对吞噬细胞杀伤的超敏性通过自主表达的HYR1而被完全补偿,而非被Bcr1p正性调节的编码GPI蛋白的其他基因。因此,HYR1是在吞噬细胞杀伤抗性途径中BCR1的下游基因。
有趣的是,HL-60来源的中性粒细胞能够在吞噬细胞和念珠菌之间的初始接触期间抑制野生型白色念珠菌中HYR1的表达。因此,在宿主吞噬细胞和白色念珠菌之间发生动态的相互作用,其中吞噬细胞介导野生型真菌中吞噬细胞抗性基因的表达延迟。这与以前的发现一致,以前发现人中性粒细胞延缓白色念珠菌菌丝的形成和菌丝共表达基因的表达[18]。
对吞噬细胞杀伤的抗性是一种复杂的表型,可能归因于多种因素。吞噬细胞可以在细胞内或细胞外杀伤念珠菌。最近,白色念珠菌细胞表面超氧化物歧化酶被表征为毒力因子,其通过降解宿主来源的高度活性氧物质而有助于真菌逃脱杀伤[22]。中性粒细胞典型地粘附菌丝形式的真菌的表面并且覆盖在其表面,因为伸展的菌丝太大使得吞噬细胞不能将其完全摄入。由于白色念珠菌在菌丝形式中表达HYR1,因此有可能Hyr1p通过阻止与吞噬细胞的表面接触来促成对吞噬细胞杀伤的抵抗力。可选地,Hyr1p可能干扰吞噬细胞的氧化或非氧化杀伤机制。
HYR1过表达的体外表型在体内得到重现。在鼠播散性感染期间,与野生型菌株相比,在具有常驻吞噬细胞的器官中HYR1的过表达导致组织真菌载量的显著增加,且HYR1的抑制导致组织真菌载量的显著减少。在肾中缺少表型可能反映了这样的事实,即肾没有常驻吞噬细胞,并且在致死性播散性念珠菌病期间流入至肾中的中性粒细胞直至感染的>24hr时才开始出现[15]。然而,用rHyr1p-N免疫接种导致针对血行播散性念珠菌病的相当大的保护作用。对于rHyr1p-N疫苗可见到的效力大于以前在用rAls1p-N和rAls3p-N疫苗免疫接种的小鼠中所见到的效力。因此,rHyr1p-N是用于播散性念珠菌病的有前途的疫苗候选物。
我们的数据还证明使用条件性过表达/抑制方式来研究指定基因的潜在毒力功能的优势。在体外研究毒力基因的大规模正向遗传方法需要功能筛选测定,并且限于筛选在进行测定时表达的基因。当在用于体外筛选测定的条件下在野生型菌株中基因不显著表达时,基因的强制过表达仍能够在测定中检测功能获得(gain-of-function)现象,如在念珠菌芽生孢子生长期间HYR1的情况下那样。当基因强有力地表达时,基因的条件性抑制在相同的测定中产生了功能缺失表型。此外,当同时使用过表达和抑制时,所述表型可以被放大,如在体内HYR1对肝和脾真菌载量的作用那样。通过比较基因过表达和抑制来检测表型的能力使得能够进行条件性基因表达以克服功能余度(functional redundancy)的限制,当研究基因家族的成员时,这种功能余度的限制尤其不利于正向遗传方法。
总之,我们使用了条件性基因表达方法来将Hyr1p鉴定为一种白色念珠菌的表面表达的毒力因子。HYR1表达在体外介导对中性粒细胞杀伤的抵抗力,并且在体内增加组织真菌载量。最后,我们证明HYR1是一种有前途的疫苗靶标,其值得进一步开发为用于播散性念珠菌病的预防策略。
材料和方法
使用下列的材料和方法获得上述的结果。
菌株和培养条件
使用的所有菌株列举在表1中,并且如前所述生长[8]。
条件性HYR1过表达/抑制突变体构建
为了产生条件性HYR1表达菌株,将HIS1-TR启动子盒[8]插入至THE4株的HYR1基因的一个等位基因前方,得到CAAH株。CAAH株中HIS1基因座处的URA3被环出,产生CAAH-1。CAAH-1中的HYR1的第二个等位基因被可循环的URA3盒破坏,产生CAAH-2株,然后将URA3环出,得到CAAH-3株。将3.9-kb含有URA3-IRO1基因的Nhe I-Pst I片段插入至CAAH-3基因组上的其原始基因座中,得到CAAH-31。使用的引物列举在表1中。
半定量RT-PCR
在体外检测基因表达的半定量RT-PCR以前已经进行了描述。用来检测EFB1表达的引物为EFB1a和EFB1b;用来扩增HYR1的引物为HYR1specific1和HYR1 specific2(表1)。为了研究中性粒细胞对白色念珠菌HYR1表达的影响,在YPD上生长过夜的1x106个SC5314细胞在添加了10%混合人血清的RPMI1640中与1x107个HL-60来源的中性粒细胞共培养或单独培养。以30min间隔取样持续3h直至提取RNA,并进行半定量RT-PCR。
吞噬细胞杀伤测定
分离人中性粒细胞,HL-60细胞分化成中性粒细胞或巨噬细胞,并且如前所述进行吞噬细胞杀伤测定[8-10]。简言之,吞噬细胞与真菌孵育1h,然后超声作用并定量培养。通过将与吞噬细胞共孵育后的真菌菌落数除以与不含吞噬细胞的培养基孵育的真菌菌落数,计算杀伤百分比。人中性粒细胞和HL-60来源的中性粒细胞或巨噬细胞分别以2∶1和20∶1的巨噬细胞:真菌比测试。对于bcr1和相关突变体,芽生孢子在添加了10%FBS的RPMI中在37℃下预先出芽40min,然后进行测定。
在光滑念珠菌BG14中HYR1的异源表达
光滑念珠菌BG14用HYR1表达载体pGRB2.2-HYR1或空载对照质粒pGRB2.2转化[11]。HYR1编码序列由CG-Hyr1-a和CG-Hyr1-b扩增(表1)并使用In-FusionTM 2.0Dry-Down PCR克隆试剂盒(Clontech Laboratories,Mountain View,CA)根据生产商的说明书克隆进pGRB2.2的Xba I,Xho I位点中。
在血原性感染期间的白色念珠菌HYR1表达
如所述的那样在血原性播散性念珠菌病期间检验野生型白色念珠菌SC5314的HYR1表达。在感染后6和24h采集BALB/C小鼠的脑、肝、肺、肾和脾。使用的引物列举在表1中。使用RETROscript(Ambion,Texas)进行逆转录。对于小鼠G3PDH看家基因的扩增,使用引物G3PDHF和G3PDHR。对于白色念珠菌的HYR1和EFB1的检测,进行两轮PCR。第一轮使用外引物组(EFB1F和EFB1R用于EFB1,或P2和P5用于HYR1);第二轮使用试样量(1μl)的第一轮PCR产物作为模板。内引物组如下:EFB1nF和EFB1nR(用于EFB1),或P2和P4(用于HYR1)。所有PCR条件如下:在95℃下变性,2min和在94℃,30s(变性),55℃,30s(复性)和72℃,90s(延伸)扩增35个循环。对于qRT-PCR,如上制备cDNA。如所述的那样优化扩增效率和实时RT-PCR SYBR green测定[12]。组成型表达的ACT1用作所有反应的对照。如在ABI PRISM 7000 Sequence Detection System User Bulletin 2(Applied Biosystems,USA)中所述进行计算和统计分析。
组织真菌载量
在从相对于感染-3天开始的整个实验时间期间给予小鼠含溶解在5%蔗糖溶液中的DOX(2mg/ml)或不含DOX的水[13],并且小鼠自由进食和饮水。如前所述进行组织真菌负载[8],不同之处在于感染后1天取出器官。所有涉及小鼠的程序遵循NIH指导原则,均由机构动物管理和使用委员会批准。
rHyr1p-N制备
rHyr1p-N(来自Hyr1p的氨基酸25-350)在大肠埃希菌(Escherichia coli)pQE-32表达系统(Qiagen)中产生,且如其他地方所述纯化6XHis标签蛋白[14],例外是使用HisPur Cobalt树脂(Thermo Scientific)亲和柱。使用Detoxi-Gel内毒素清除柱(Endotoxin Removing Columns)(Thermo Scientific)从rHyr1p-N中去除内毒素,并且根据生产商的说明书用Limulus Amebocyte Lysate endochrome(Charles River)测定内毒素水平。使用此程序,内毒素减少至用于接种的每个剂量<0.28EU。
Hyr1p细胞定位的免疫荧光检测
使用通过用rHyr1p-N(来自氨基酸25-350)免疫接种小鼠产生的多克隆抗-Hyr1p抗血清进行间接免疫荧光。hyr1缺失菌株的1x107个芽生孢子接种物在RPMI 1640中在37℃下孵育90min并离心两次以吸收抗血清。
白色念珠菌芽生孢子(1x105)在RPMI 1640中在37℃下预先出芽90min,并转移至4-孔室玻片(Nalge Nunc International Corp,IL,USA)中。在4℃下孵育30min后,细胞用300μl 1.5%山羊血清封闭,用1∶100稀释度的多克隆抗血清或作为阴性对照的PBS、然后通过1∶200的异硫氰酸荧光素标记的山羊抗鼠IgG染色。通过共聚焦扫描激光显微镜对细胞成像[15]。
免疫接种方案
所有免疫接种在颈根部皮下进行。8只幼年(10-12-周)C57BL/6小鼠用20μg完全弗氏佐剂中的亲和纯化的rHyr1p-N接种,并在3周时用不完全弗氏佐剂(IFA)中的亲和纯化的rHyr1p-N强化免疫。另外8只幼年小鼠接受与由用空载质粒转化的大肠杆菌细胞产生的制剂混合的单独佐剂。强化免疫后14天,小鼠经尾静脉用5x105个野生型白色念珠菌SC5314细胞感染[16]。
还使用明矾(2%铝胶(Alhydrogel);Brenntag Biosector)评价rHyr1p-N在对抗血原性播散性念珠菌病中的效力,明矾是由食品药品监督管理局(FDA)批准用于人类的一种佐剂。另外,为确定rHyr1p-N对白色念珠菌的其他菌株有保护作用,我们使用了另一临床分离株,15563株。对于这些实验,33μg亲和纯化的rHyr1p-N与0.1%铝胶混合并如上在第0天施用于BALB/c小鼠,在第21天强化,然后在第35天通过尾静脉注射用白色念珠菌感染。对于所有接种实验,感染后35天小鼠存活用作终点。
F(ab)’2阻断测定。从用rHyr1p-N或由用空载质粒转化的大肠杆菌细胞产生的制剂接种的5只小鼠收集混合的抗-Hyr1p或对照血清。使用Nab Spin Kit(Thermo Scientific)分离来自两种血清的总IgG。F(ab)’2片段用Pierce F(ab)’2制备试剂盒根据生产商的说明书纯化。念珠菌细胞在冰上用5%正常小鼠血清(Santa Cruz Biotechnology)或添加了由rHyr1p-N免疫小鼠或对照小鼠IgG制备的5%F(ab)’2的5%正常小鼠血清调理45min,然后与小鼠中性粒细胞混合。小鼠中性粒细胞杀伤测定如上所述。
统计学分析
根据情况,不同组之间吞噬细胞介导的杀伤和组织真菌载量通过Mann-Whitney U检验进行未配对比较。非参数对数秩检验(non-parametric Log Rank test)用来确定生存时间的差异。P值<0.05被认为是有显著性。
表1在本研究中使用的菌株和寡核苷酸
菌株
寡核苷酸
实施例II
rHyr1p-N是对抗播散性念珠菌病的疫苗
背景:我们已经发现白色念珠菌过表达HYR1介导了体外对中性粒细胞杀伤的抵抗力。我们力图确定在感染期间HYR1过表达对体内组织真菌载量的影响,并且确定用rHyr1p-N免疫接种保护小鼠免受播散性念珠菌病的可能性。
方法:小鼠经尾静脉用HYR1过表达/抑制或野生型白色念珠菌感染。在感染后1天收获肝和脾,并且通过qRT-PCR和定量培养分别测定HYR1的表达水平和组织真菌载量。对于接种,rHyr1p-N在大肠杆菌pQE-32表达系统中产生,并根据生产商(Qiagen)的说明书纯化。小鼠用20μg完全弗氏佐剂(CFA)中的rHyr1p-N接种,并在3周时用不完全弗氏佐剂(IFA)中的rHyr1p-N强化免疫,并在强化免疫后2周用白色念珠菌SC5314株感染。对照小鼠接受佐剂结合来自用空载质粒转化的大肠杆菌的细胞提取物。
结果:与对照菌株相比,HYR1的过表达显著地增加了肝和脾中的真菌载量。与对照菌株相比,HYR1的抑制显著地减少了肝和脾中的真菌载量。在用过表达或抑制菌株相比于用对照菌株感染的肝中HYR1的相对表达水平分别为2.5和0.8。rHyr1p-N疫苗导致感染小鼠的长期存活率为62.5%,相比在对照小鼠中存活率为0%。
结论:HYR1表达影响白色念珠菌在体内感染组织的能力。此外,用rHyr1p-N接种明显地保护小鼠免受播散性念珠菌病。rHyr1p-N疫苗可用于预防播散性念珠菌病。
参考文献
1.Spellberg BJ,Filler SG,and Edwards JE,Jr.Current treatment strategies for disseminated candidiasis.Clin Infect Dis.2006;42:244-251
2.Del Poeta M.Role of Phagocytosis in the Virulence of Cryptococcus neoformans.Eukaryot Cell 2004;3:1067-1075
3.Koh AY,Kohler JR,Coggshall KT,Van Rooijen N and Pier GB.Mucosal damage and neutropenia are required for Candida albicans dissemination.PLoS Pathog.2008;4:DOI:10.1371/journal.ppat.0040035.
4.Gulay Z,Imir T.Anti-candidial activity of natural killer(NK)and lymphokine activated killer(LAK)lymphocytes in vitro.Immunobiology 1996;195:220-230
5.Stone HH.Studies in the pathogenesis,diagnosis,and treatment of Candida sepsis in children.J Pediatr Surg.1974;9:127-133
6.Richard M,Ibata-Ombetta S,Dromer F,Bordon-Pallier F,Jouault T and Gaillardin C.Complete glycosylphosphatidylinositol anchors are required in Candida albicans for full morphogenesis,virulence and resistance to macrophages.Mol.Microbiol.2002;44
7.Bailey DA,Feldmann PJ,Bovey M,Gow NA and Brown AJ.The Candida albicans HYR1 gene,which is activated in response to hyphal development,belongs to a gene family encoding yeast cell wall proteins.J Bacteriol.1996;178:5353-5360
8.Fu Y,Luo G.,Spellberg BJ,Edwards JE,Jr,and Ibrahim AS.Gene overexpression/suppression analysis of candidate virulence factors of Candida albicans.Eukaryot Cell.2008;7:483-492
9.Nusing R,Goerig M,Habenicht AJ and Ullrich V.Selective eicosanoid formation during HL-60 macrophage differentiation.Regulation of thromboxane synthase.Eur J Biochem 1993;212:371-376
10.Spellberg BJ,Collins M,French SW,Edwards JE,Jr.,Fu Y and Ibrahim AS.A phagocytic cell line markedly improves survival of infected neutropenic mice.J Leukoc Biol 2005;78:338-344
11.Eiden-Plach A,Zagorc T,Heintel T,Carius Y,Breinig F.Viral preprotoxinsignal sequence allows efficient secretion of green fluorescent protein by Candida glabrata,Pichia pastoris,Saccharomyces cerevsiae,and Schizosaccharomyces pombe.Appl.Environ.Microbiol 2004;70:961-966
12.Avrova AO,Venter E,Birch PR and Whisson SC.Profiling and quantifying differential gene transcription in Phytophthora infestans prior to and during the early stages of potato infection.Fungal Genetics & Biology 2003;40:4-14
13.Saville SP,Lazzell AL,Monteagudo C and Lopez-Ribot JL.Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection.Eukaryot Cell 2003;2:1053-1060.
14.Spellberg B,Ibrahim AS,Yeaman MR,et al.The antifungal vaccine derived from the recombinant N terminus of Als 3p protects mice against the bacterium Staphylococcus aureus.Infect Immun.2008;76:4574-4580
15.Fu Y,Ibrahim AS,Sheppard DC,Chen YC,French SW and Cutler JEea.Candida albicans Als 1p:an adhesin that is a downstream effector of the EFG1fiamentation pathway.Mol Microbiol 2002;44:61-72
16.Ibrahim AS,Spellberg BJ,Avenissian V,Fu Y,Filler SG and Edwards JEJ.Vaccination with rAls1p-N improves survival during murine disseminated candidiasis by enhancing cell-mediated,not humoral,immunity.Infect Immun2005;73:999-1005
17.Nobile CJ,Andes DR,Nett JE,et al.Critical role of Bcrl-dependent adhesins in C.albicans biofilm formation in vitro and in vivo.PLoS Pathog.2006;2:e63
18.Fradin C,De Groot P,MacCallum D,et al.Granulocytes govern the transcriptional response,morphology and proliferation of Candida abicans in human blood.Molecular Microbiology 2005;56:397-415
19.Castano I,Pan SJ,Zupancic M,Hennequin C,Dujon B and Cormack BP.Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata.Mol Microbiol 2005;55:1246-1258
20.Schofield DA,Westwater C,Warner T,Nicholas PJ,Paulling EE and Balish E.Hydrolytic gene expression during oroesophageal and gastric candidiasis inimmunocompetent and immunodeficient gnotobiotic mice.J Infect Dis2003;188:591-599
21.Kumamoto CA,Vinces MD.Contributions of hyphae and hypha-co-regulated genes to Candida albicans virulence.Cell Microbiol.2005;7:1546-1554
22.Frohner IE,Bourgeois C,Yatsyk K,Majer O and Kuchler K.Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance.Mol Microbiol 2009;71:240-252
作为针对念珠菌的主动和被动免疫接种的靶标的HYR1专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。
















动态评分
0.0