











专利摘要
本发明公开了一种新的三萜合成酶的功能及应用。本发明提供的0s0SC8蛋白在制备三萜化合物中的应用;所述0s0SC8蛋白的氨基酸序列为序列表中的序列2。本发明的实验证明,本发明提供水稻中0s0SC8基因的生化功能为合成(3S,13S)-malabarica-17,21-dien-3β,14-diol,从而证明,0s0SC8基因编码(3S,13S)-malabarica-17,21-dien-3β,14-diol合成酶。
说明书
技术领域
本发明涉及生物技术领域,尤其涉及一种新的三萜合成酶的功能及应用。
背景技术
三萜化合物包括游离三萜和三萜皂苷,广泛存在于高等植物中。三萜化合物是通过异戊二烯途径来合成的,其生物合成路线为:甲羟戊酸(Mevalonic acid)生成的异戊烯二磷酸(IPP)及其异构体二甲基丙烯二磷酸(DAPP)在香草二磷酸合成酶(GPS)作用下首先形成香叶二磷酸(GPP),接着利用法呢二磷酸合成酶(FPS)转化成法呢二磷酸(FPP),又在鲨烯合成酶(Squalene synthase,SQS)的作用下合成鲨烯,然后经鲨烯环氧酶(Squalene epoxidase,SQE)催化转变为2,3-氧化鲨烯(2,3-oxidosqualene)。2,3-氧化鲨烯在氧化鲨烯环化酶(OSCs)的作用下环化形成三萜化合物的前体物质,如β-香树素(β-amyrin)。目前已经从拟南芥、水稻、苜蓿、百脉根等不同的植物中分离得到了几十个编码OSCs酶的基因,除了合成植物甾醇的环阿屯醇合成酶(cycloartenol synthase,CAS),还包括arabidiol synthase,羽扇醇合成酶(lupeol synthas,LUP)、β-香树素合成酶(β-Amyrin synthase,βAS)、葫芦二烯醇合成酶(cucurbitadienol,CPQ)、thalianol synthase(THA)、marneral synthase(MRN)、camelliol synthase(CAMS)、baruol synthase(BARS)、isomultiflorenol synthase(IMS)等。另外,人们从双子叶植物中发现了羊毛甾醇合成酶(lanosterol synthase,LAS),并证实在拟南芥中参与了植物甾醇的合成。三萜化合物骨架在依赖细胞色素P450单加氧酶、糖基转移酶和酰基转移酶进行氧化、糖基化及酰基化等化学修饰,最终获得不同种类的三萜皂苷。
众所周知,三萜化合物是一种重要的天然药物,例如,齐墩果酸具有降低转氨酶的功能,临床用于治疗急性黄疸肝炎;人参皂苷具有增加白细胞数量、提高人体免疫力、促进物质代谢、抗疲劳、抗衰老等作用。但三萜化合物的生物学功能的了解还非常少,例如燕麦在根尖合成的燕麦苷(avenacins)具有广谱的抗菌功能,能防止多种病原菌侵染根部,发挥抗病作用。
在水稻中发现有7个可编码全长OSCs酶的基因序列,并鉴定了其中4个OSC基因的生化功能,它们分别是环阿屯醇合酶基因(0s02g04710/0s0SC3),帕克醇(parkeol)合酶基因(0s11g08569/0s0SC9),蓍醇B (achilleol B)合酶基因(0s11g181/0s0SC10),以及异山苷子萜醇(isoarborinol)合酶基因(0s11g35710/0s0SC12),还有一些OSC基因的功能不清楚。
发明内容
本发明的一个目的是提供0s0SC8蛋白的应用。
本发明提供了0s0SC8蛋白在制备三萜化合物中的应用;
所述0s0SC8蛋白的氨基酸序列为序列表中的序列2。
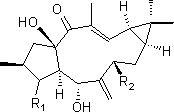
所述三萜化合物的结构式如下式I所示:
式I。
所述0s0SC8蛋白的编码基因的核苷酸序列为序列表中的序列1。
本发明的另一个目的是一种制备三萜化合物的方法。
本发明提供的方法,包括如下步骤:
1)将0s0SC8蛋白的编码基因导入目的植物得到转基因植物,
2)提取步骤1)得到转基因植物的叶片,收集提取产物,即得到三萜化合物;
所述0s0SC8蛋白的氨基酸序列为序列表中的序列2。
步骤1)中,所述蛋白的编码基因通过重组载体导入目的植物;
步骤2)中,所述提取步骤1)得到转基因植物的叶片的方法包括如下步骤:
A、用裂解液裂解所述转基因植物的叶片,得到裂解产物、过滤,收集滤液;
B、将步骤A得到的滤液用等体积正己烷萃取,收集正己烷层,再用等体积饱和NaCl水溶液洗涤,收集正己烷层,即得到三萜化合物。
步骤1)中,所述0s0SC8蛋白的编码基因的核苷酸序列为序列表中得序列1;
所述重组载体为将所述0s0SC8蛋白的编码基因插入表达载体pH7WG2D,1,得到表达所述蛋白的载体;
步骤2)中,所述裂解液按照如下方法制备:将KOH、乙醇和水混合,得到裂解液,所述KOH在所述裂解液中的浓度为10%(质量百分含量),所述乙醇(EtOH)在所述裂解液中的浓度为80%(体积百分含量);
所述裂解液和所述转基因植物的叶片的配比为5L∶500g;
所述裂解的温度为80℃,所述裂解的时间为2h;
所述三萜化合物的结构式如下式I所示:
式I。
所述目的植物为单子叶植物,所述单子叶植物为水稻,具体为粳稻品种中花11号。
本发明的第三个目的是提供0s0SC8蛋白的应用。
本发明提供了0s0SC8蛋白作为三萜化合物合成酶中的应用;
所述0s0SC8蛋白的氨基酸序列为序列表中的序列2;
所述三萜化合物的结构式如下式I所示:
式I。
本发明的第四个目的是提供一种重组载体。
本发明提供的重组载体,为将所述0s0SC8蛋白的编码基因插入表达载体pH7WG2D,1,得到表达所述蛋白的载体。
所述0s0SC8蛋白的编码基因的核苷酸序列为序列表中的序列1。
本发明的实验证明,本发明提供水稻中0s0SC8基因的生化功能为合成(3S,13S)-malabari ca-17,21-dien-3β,14-diol,从而证明,0s0SC8基因编码(3S,13S)-malabarica-17,21-dien-3β,14-diol合成酶。
附图说明
图1为RT-PCR的琼脂糖电泳图
图2为pCR8/GW/TOPO结构示意图
图3为pH7WG2D,1-0s0SC8结构示意图
图4为转0s0SC8水稻的western blot分析结果
图5为TLC检测结果
图6为GCMS检测结果
图7为质谱分析结果
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1、0s0SC8基因的获得
根据水稻基因组序列,设计0s0SC8的引物:5’端引物:5`-ATGTGGAAGCTCAAGATTGCCGAG-3`;3’端引物:5`-TCAAGTTGGCGCTGTTGTACTTGC-3`。以粳稻品种中花11号(Oryza sativa L.spp.Japonica,水稻花培育种前景,李梅芳,作物杂志03:17-18,1991,公众可从中国科学院植物研究所获得。)花总RNA为模板,在上述引物对的引导下,RT-PCR扩增0s0SC8的cDNA序列,具体方法包括以下步骤:
1、总RNA的提取:用Trizol法(所用试剂购自Invitrogen公司)提取2周幼苗总RNA,具体方法为:收集水稻材料100mg,立即置于液氮中研磨,加入1mL Trizol试剂,充分混匀后,25℃放置5分钟;加入0.2mL氯仿,剧烈振摇15秒,25℃温育3分钟;4℃,12000g离心15分钟;将上清液转移到一个新的1.5mL离心管中,加入0.5mL异丙醇沉淀RNA;最后将RNA沉淀用1mL 75%乙醇洗涤后溶于适量经DEPC处理过的双蒸水中,-70℃保存备用。
2、第一链cDNA的合成:用Superscript II RT试剂盒(Invitrogen)并按试剂盒说明书进行操作:取5μg步骤1获得的水稻总RNA放入灭活了RNase的PCR管中,加入Oligo(dT)12-18(500mg/mL)1μL和dNTP Mix(10mM each)1μL,用DEPC处理后的双蒸水补充至12μL,混匀后在65℃下加热5分钟,然后迅速置于冰上,1分钟。短暂离心后再加入5×第一链合成缓冲液4μL、0.1M DTT 2μL和RNaseOut(40unit/μL)1μL,轻轻混匀后,42℃温育2分钟,然后加入Superscript II反转录酶(200unit/μL)1μL,混匀,42℃温育50分钟,70℃加热15分钟使酶失活,得到第一链cDNA。
3、0s0SC8cDNA的合成:取1μL步骤2获得的反转录产物cDNA为模板,在5’端引物和3’端引物的引导下,进行PCR,PCR反应体系为:Platinum(invitrogen公司)0.5μL、10×缓冲液5μL、dNTP 1μL、5’端引物(10μM/L)1μL、3’端引物(10μM/L)1μL、模板1μL、加双蒸水补充反应体系至50μL。PCR反应条件为:先95℃5分钟;再94℃45秒,60℃45秒,72℃5分钟,共35个循环;最后72℃10分钟。
反应结束后,对PCR产物进行0.8%琼脂糖凝胶电泳检测,检测结果如图1所示(泳道D15000为DNA标准分子量D15000,泳道1-5为RT-PCR产物),得到分子量约为2.3kb的条带,与预期结果相符。
用琼脂糖凝胶回收试剂盒(北京天为时代公司)回收该片段,然后将该回收片段与载体pCR8/GW/TOPO(Invitrogen,K250020,结构示意图如图2)进行连接,连接体系为:PCR产物4μL,salt solution 1ul TOPO vector 1μL,25℃反应20min。参照Cohen等的方法,将连接产物转化大肠杆菌DH5α感受态细胞,得到重组子,根据pCR8/GW/TOPO载体上的壮观霉素抗性标记筛选阳性克隆。提取阳性克隆的质粒,送去测序,以该质粒载体上的M13F和M13R序列为引物对其进行核苷酸序列测定,测序结果表明该质粒连接的PCR产物具有序列表中的序列1所示的核苷酸,该PCR产物即为基因0s0SC8,编码的蛋白命名为0s0SC8,其氨基酸序列为序列表中的序列2。该质粒为将序列表中序列1所示的核苷酸插入pCR8/GW/TOPO中得到的质粒,命名为pCR8-0s0SC8。
序列表中的序列1由2358个碱基组成,其开放阅读框(ORF)为序列1自5’端第1-2358位碱基。序列2由785个氨基酸组成。
实施例2、转0s0SC8水稻获得及0s0SC8的功能研究
一、转0s0SC8水稻获得
1、0s0SC8物表达载体的构建
1)用限制性内切酶Mlu I将pCR8-0s0SC8完全酶切。酶切体系为:5μg质粒、2.5μL 10×酶切缓冲液、1μL Mlu I,加ddH2O补充反应体系至50μL。酶切反应条件为:37℃酶切1小时。
2)用琼脂糖电泳对酶切产物进行分离,回收线性化的产物,溶解于20μL ddH2O中。
3)将步骤2)获得的线性化的产物进行LR反应。5μl反应体系中加下列组分:0.5μL终端载体pH7WG2D,1(Karimi,M.,Inzé,D.and Depicker,A.(2002)GATEWAYvectors for Agrobacterium-mediated plant transformation,Trends in Plant Science;7(5):193-5,公众可从中国科学院植物研究所获得。)(200ng/μl)、2μL线性化产物(50ng/μL)、1μL LR mix、1.5μl TE pH8.0,25℃温育4h。加0.5μLproteinase K,37℃,10min。
4)将LR反应的产物转化大肠杆菌DH5α感受态细胞,用含有壮观霉素的LB平板(spe浓度为100ug/ml)进行筛选,得到阳性克隆。
5)提取阳性克隆的质粒进行PCR鉴定,引物如下:5`-ATGTGGAAGCTCAAGATTGCCGAG-3`;5`-TCAAGTTGGCGCTGTTGTACTTGC-3`,结果表明,该质粒为将序列表中的序列1重组到pH7WG2D,1,将该重组质粒命名为pH7WG2D,1-0s0SC8(见图3)。
2、将pH7WG2D,1-0s0SC8导入农杆菌(EHA105)
1)冰浴电击杯,取50ng质粒pH7WG2D,1-0s0SC8于100μl农杆菌EHA105(A.tumefaciens,Hood,E.E.,Gelvin,S.B.,Melchers,L.S.& Hoekema,A.(1993).New Agrobacterium helper plasmids for gene transferto plants.Transgen.Res.2,208-218.公众可从中国科学院植物研究所获得。)感受态细胞,轻轻混匀,转入1mm电击杯(Bio-RAD),盖上盖。
2)1800V电击(德国eppendorf公司),迅速加入900μl YEB培养基,混匀,28℃,200rpm摇1h。
3)将菌液涂于含有50ug/ml利福平和100ug/ml壮观霉素的YEB固体培养基板上,28℃,两天,得到阳性克隆,提取阳性克隆的质粒送去测序,结果为该质粒为pH7WG2D,1-0s0SC8,将该阳性克隆命名为EHA105/pH7WG2D,1-0s0SC8。
3、将0s0SC8转入野生型水稻
1)在含2mg/L的2,4-D的N6培养基上诱导成熟胚的水稻(粳稻品种中花11号,以下简称为野生型水稻,)愈伤组织。
2)将EHA105/pH7WG2D,1-0s0SC8在28℃,200rpm培养到OD600=1.0。3000rpm离心收集菌体,AAM培养基悬浮。
3)把愈伤组织在农杆菌悬浮液中浸泡30分钟,在滤纸上吸干,置于含乙酰丁香统(AS)的N6培养基上共培养3天。
4)用头孢霉素(cefotaxime)和羧苄青霉素(carbenicillin)洗去多余农杆菌,沥干。
5)置于25mg/L的hygromycin B的N6筛选培养基上筛选2周,提高hygromycinB浓度(50mg/L)继续筛选2周,
6)在包含NAA和6-BA的再生MS培养基上(16小时白昼22℃/8小时黑夜18℃)长苗,获得16株T0代转0s0SC8水稻,移栽到温室(12小时白昼30-35℃/12小时黑夜18~22℃)中生长。
4、0s0SC8过表达株系外源基因的表达验证
提取T0代转0s0SC8水稻植株叶片的总蛋白,进行western blot分析,表明0s0SC8在部分水稻株系中成功过量表达。
1)植株叶片总蛋白的提取:
剪取自然条件网室生长的生殖期(8-12周)T0代转0s0SC8水稻水稻叶片,在液氮中研磨成粉末状,用预冷的小勺将粉末装入1.5ml离心管中,每克粉末加入3ml水稻总蛋白抽提液,立即涡旋混匀,冰浴10min;8000rpm离心15min,收集蛋白上清液于新的1.5ml离心管中,加入等体积的2×SDS loading buffer,用手指轻弹混匀后,沸水浴10min。将变性的蛋白质在室温(25℃)下8000rpm离心5min,使杂质沉入管底,4℃保存备用。
水稻总蛋白抽提液配方:
2)PAGE胶电泳:
制备8%的PAGE胶,每份叶片总蛋白上样5μl,蛋白预染Marker 5μl,100V电泳10min后150V电泳1h。
3)转膜:
切取分离胶,量取长宽后,在转移缓冲液中浸泡30min;剪取同分离胶大小一样的PVDF膜,甲醇中浸泡5min后,置于转移缓冲液中浸泡15min。同时剪取和分离胶大小一样的两张滤纸,浸泡于转移缓冲液中;将海绵垫浸湿后放在转移孔板黑面上,再依次放上滤纸、分离胶、PVDF膜、滤纸、海绵垫,注意赶走各层之间的气泡,然后合上转移孔板的黑白面并夹紧;把转移芯放在电泳槽中,并加入大半槽的转膜缓冲液,按照转移孔板的黑面对应转移芯黑色面,红面对应红色面的方法,将夹板插于转移芯中,补加转膜缓冲液至blotting刻度线。按红色电极对应红线,黑色电极对应黑线的方法盖上电泳槽盖后,接上电泳仪,然后把电泳槽放于4℃冰箱中,100V转膜1h20’。
4)封闭和抗体孵育:
将膜浸于TBST缓冲液配制的5%脱脂牛奶中,4℃封闭过夜。
一抗:从脱脂牛奶中取出PVDF膜,用TBST洗去表面的牛奶后,滴去表面多余的缓冲液后,正面朝上(蛋白与PVDF膜结合的那一面)放在parafilm封口膜上(封口膜平铺于培养皿底),将抗0s0SC8的特异性抗体13G12(由Abmart公司制备,本抗体是定制,通过合成多肽免疫小鼠细胞系获得,多肽片段是KLKIAEGGPWLKSGNSHVGRETWEFDPNFGTSEEREAVEAARIEFQKNRFRTRHTSDVLARMQLAKANNFSIDLQKEKDGNPINIDTATVSDILKKALSYFSAIQAYDGHWPGDFPGPLFTTATMIIVLYVTESLTITLSSEHHKEICRYLYNRQNIDGGWGLHAEGESSMLSTALNYTALRLLGENVDDGPDISMHKARK)按1∶500的比例用TBST缓冲液配制的1%脱脂牛奶稀释后,用移液枪滴于整张PVDF膜上,盖上培养皿盖,室温静止杂交1h。
二抗:先用TBST洗去膜表面的杂交液后,再在摇床上用TBST洗膜3次,每次10min。滴去表面多余的缓冲液后,正面朝上放在parafilm封口膜上,将康为世纪公司生产的HRP标记的山羊抗小鼠(CW0122)的二抗按1∶5000的比例用TBST缓冲液配制的1%脱脂牛奶稀释后,用移液枪滴于整张PVDF膜上,盖上培养皿盖,室温静止杂交1h。
4)化学发光和显影:
先用TBST洗去膜表面的杂交液后,再用TBST洗膜3次,每次10min。在平整的桌面上铺一张长宽约20cm的保鲜膜,用吸水纸吸去PVDF膜表面的液体后,正面朝上,平铺于保鲜膜上。将Thermo scientific公司生产的SuperSignal West Dura Extended Duration Substrate化学发光试剂盒中的A、B液按1∶1混合后,滴于膜的表面,静置5min。打开暗盒,剪一张长宽比暗盒盖子及底部总大小稍小的保鲜膜,铺于暗盒中,盒底部分的保鲜膜用两滴水轻轻的贴在底部。弃去膜表面的发光液,用吸水纸吸去PVDF膜表面多余液体,将PVDF膜正面朝上并且按上下方向正确地平铺于暗盒底部的保鲜膜上,再盖上盒盖部分的保鲜膜,注意盒盖部分的保鲜膜与PVDF膜之间不要产生气泡,盖好后关灯。
取出柯达X-film胶片,在右下角剪去一个三角形的角(以示方向),观察好荧光情况后迅速准确地压在暗盒中,2s-10s(视情况而定,如果荧光强,2s即可,如果弱,可能需要1min以上)后迅速取出胶片,置于显影液中,直到条带显出,取出胶片放入水中清洗几下后,滴去多余的水后放入定影液中,2-3min后开灯观察显影效果。从定影液中取出胶片,用水清洗后晾干,观察记录后扫描保存。
以野生型水稻的花和叶片为对照。
T0代转0s0SC8水稻的western blot分析结果如图4所示,其中,WT F为野生型水稻花,WT L为野生型水稻叶片,OE8-4-OE8-10:为编号为OE8-4-OE8-10的T0代转0s0SC8水稻叶片,可以看出,野生型水稻叶片中没有目的蛋白,野生型水稻花中有目的蛋白,编号为OE8-4、OE8-5、OE8-7、OE8-8、OE8-9、OE8-10的T0代转0s0SC8水稻的叶片中有88.8KD的蛋白表达,与预期结果一致,为阳性T0代转0s0SC8水稻。
收集阳性T0代转0s0SC8水稻种子、播种、传代,直到得到T3代转0s0SC8水稻.
采用同样的方法,将空载体pH7WG2D,1转入野生型水稻中,得到转空载体水稻,同样westenblot检测,结果转空载体水稻的叶片仍然没有目的蛋白,说明得到转空载体水稻。
二、0s0SC8的功能研究
利用OE8过表达水稻株系产生(3S,13S)-malabarica-17,21-dien-3β,14-diol:
1)称编号为OE8-10的T3代转0s0SC8水稻的叶片和野生型水稻的叶片,分别约500g,液氮磨样,加入碱裂解液5L(10%KOH(w/v),80EtOH(乙醇,v/v),所述裂解液按照如下方法制备:将KOH、EtOH和水混合,得到裂解液,所述KOH在所述裂解液中的浓度为10%(质量百分含量),所述EtOH在所述裂解液中的浓度为80%(体积百分含量),80℃水浴2h,其间不断混匀样品。
2)过滤去掉沉淀,等体积正己烷萃取三次,等体积饱和NaCl水溶液洗三次,合并正己烷层,减压蒸馏,分别得到编号为OE8-10的T3代转0s0SC8水稻粗提物、野生型水稻粗提物。
3)乙醚溶粗提物样品,少量样品TLC检测:展开剂:正己烷∶乙醚=1∶1;显色剂:2%(体积百分含量)茴香醛和2%(体积百分含量)硫酸乙酸,125℃下显色。
以转空载体水稻为对照。
将上述步骤3)得到的粗提物进行TLC检测结果见图5,OE8为编号为OE8-10的T3代转0s0SC8水稻,Z11为野生型水稻,从图中看出,编号为OE8-10的T3代转0s0SC8水稻粗提物比野生型水稻粗提物多个斑点(箭头所指),即可能多一种化合物。
将上述步骤3)得到的粗提物进行GCMS分析,GCMS分析方法:(安捷伦7890,进样1ul;氦气流速:0.8ml./min;GC方法:柱子DB-5,进样口280℃,转移线280℃,升温程序60-220℃30℃/min,220℃保持3min,220-300℃2℃/min,300℃保持5min;MS方法:50-600MS,检测器电压1600v,电子能量-70V),结果如图6所示,上图表示编号为OE8-10的T3代转0s0SC8水稻,下图表示野生型水稻,编号为OE8-10的T3代转0s0SC8水稻比野生型水稻多一个目的峰(箭头所指),该目的峰对应的保留时间为43.742min,该峰的化合物的分子量为426.4。
转空载体水稻和野生型水稻的结果无显著差异。
通过上述结果看出,编号为OE8-10的T3代转0s0SC8水稻粗提物比野生型水稻粗提物多一个分子量为426.4的化合物。然后对编号为OE8-10的T3代转0s0SC8水稻进行大量提取,重复步骤1)-3)。
4)将上述粗提物在直径24mm层析柱中填入30g粒径200目的层析硅胶,用正己烷∶乙醚=1∶1洗脱,收集组分47-62/10ml,挥干后溶于乙醚。
5)在直径24mm层析柱中填入8g粒径200目的层析硅胶,用正己烷∶乙醚=4∶1洗脱,收集组分104-150/2ml挥干后,得到提取产物。
将提取产物进行TLC检测,结果提取产物为分子量为426.4的化合物。
6)提取3次500g叶片共得到干重14mg的提取产物/1500g叶片,取干重14mg的提取产物进行1H-、13C-NMR(400MHz)和MS分析。
提取产物的质谱分析(方法同GCMS),结果如图7所示。
提取产物的1H-、13C-NMR(北京大学核磁中心布鲁克400MHz,溶剂峰δH=7.28ppm,δC=128.0ppm作为内参),
结果如表1所示,
表1为400Hz NMR date,δppm,in C6D6
文献报道的结构数据如下(Hoshino et al.,Angew.Chem.Int.Ed.2004.43:6700-6703):EI-MS m/z:69(100),93(61),135(63),189(65),207(45),229(50),247(28),426(M+H2O,12%),具体见表2:
表2为600Hz NMR date,δppm,in C6D6
从上述可以看出,确定提取产物为(3S,13S)-malabarica-17,21-dien-3β,14-diol,为一种三萜化合物,结构式如式I所示:
式I。
一种新的三萜合成酶的功能及应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。













动态评分
0.0