






专利摘要
本发明涉及合成生物学技术领域,特别涉及基因元件、表达载体及其应用。本发明提出一种提高脂肪烃微生物合成的新方法:通过动态平衡关键中间代谢物脂肪醛的积累和转化来提高大肠杆菌脂肪烃合成的能力。为了达到这个目的,本发明首先通过协同表达脂肪醇合成过程,通过竞争路径帮助ADO平衡脂肪醛的合成与转化;随后将与脂肪醛动态平衡相关的脂肪酸合成、脂质降解和电子传递三个模块与脂肪烃合成模块相适配,最终实现工程大肠杆菌高产脂肪烃的人工合成。
权利要求
1.aar基因和ado基因共表达在提高脂肪烃产量中的应用。
2.根据权利要求1所述的应用,其特征在于,aar基因和ado基因的表达强度为(0.60-1.0):(0.20-0.4)。
3.根据权利要求1或2所述的应用,其特征在于,还包括adhP基因与所述aar基因和ado基因共表达。
4.根据权利要求3所述的应用,其特征在于,aar基因、ado基因和adhP基因的表达强度为(0.60-1.0):(0.20-0.4):(0.10-0.60)。
5.根据权利要求1至4任一项所述的应用,其特征在于,还包括过表达fabD基因、fab基因G或fabI基因中的一个或多个。
6.根据权利要求1至5任一项所述的应用,其特征在于,不包括fabH基因、fabA基因、fabZ基因、fabB基因或accBCDA基因中的一个或多个基因的过表达。
7.根据权利要求1至6任一项所述的应用,其特征在于,其还包括IPTG的诱导,所述IPTG的浓度为0.1mM。
8.根据权利要求1至7任一项所述的应用,其特征在于,还包括aasS基因或者aasS-lipA基因共表达。
9.根据权利要求1至8任一项所述的应用,其特征在于,还包括来源于蓝细菌中的Se.Fd/FNR的过表达。
10.根据权利要求1至9任一项所述的应用,其特征在于,不包括大肠杆菌内源的Ec.Fd/FNR的过表达。
11.权利要求1至10任一项所述应用获得的基因元件或表达载体。
12.根据权利要求10所述的表达载体,其特征在于,其为大肠杆菌,其发酵的方式为分批补料发酵;所述分批补料发酵为在LB固体平板上30℃过夜培养,挑重组子单菌落接种于2mL LB培养基中30℃培养至OD为2.5~4,并按照1:100的比例转接于20mL M9培养基中,30℃培养至OD为2.5~4,并按照1:100的比例再次转接于800mL M9培养基中;当OD涨至2.5~4的时候,将培养液离心浓缩至50mL,并接种于发酵罐培养基中分批补料发酵;当OD升至15的时候,用10μM的IPTG进行诱导。
13.根据权利要求11或12所述的基因元件或表达载体在提高脂肪烃产量中的应用。
说明书
技术领域
本发明涉及合成生物学技术领域,特别涉及基因元件、表达载体及其应用。
背景技术
与生物短链醇分子相比,中长链烃类物质(C8-C18)具有热值高(单位燃料体积燃烧产生的热量最高)、蒸汽压低(安全)、以及吸湿性低(方便贮存和运输)等优点,并且在结构和化学特性上都与现行的石油基燃料相类似,没有混合壁垒,因此是比短链醇更为理想的生物替代燃料。
微生物中脂肪烃的合成主要以内源的脂肪酸碳链延长为基础。2010年,LS9公司Schirmer领导的研究团队采用基因组挖掘(genome mining)的研究思路,将10种产烃蓝细菌与1种不能产生链烷烃的蓝细菌进行了基因比较,得到了一条以脂肪酸碳链延长反应的中间代谢物——脂酰-ACP为底物的烃类合成路径,即脂酰-ACP(Cn)通过酰基-ACP还原酶(Acyl-ACP Reductase,AAR)的作用生成同样链长的脂肪醛,随后经过醛基脱甲酰氧化酶(aldehyde deformylating oxygenase,ADO)的作用脱掉醛基形成Cn-1的烷烃或者烯烃。该工作还将这两个基因在大肠杆菌MG1655体内进行表达,使其具有了烃类合成的功能。
虽然微生物中已经能够成功的合成出脂肪烃,但是烃的产量还远远达不到工业应用的要求。ADO的催化活性很差是脂肪烃合成能力差的主要原因,此外,ADO的转化效率还受制于脂肪醛底物浓度。但是在细胞内脂肪醛的积累不易过高,否则可能会造成代谢紊乱或者对细胞产生毒性。这两条明显冲突的原因表明,脂肪醛是脂肪烃合成过程中的一个关键节点,平衡中间代谢物脂肪醛的含量将是一种提高脂肪烃积累的有效手段。为了实现这个目的,已经有科学家尝试利用以RNA或者DNA为材料的生物脚手架(scaffold)将AAR和ADO进行连接,以期使得一方面能够为ADO提供更高的局部脂肪醛底物浓度,另一方面使得脂肪醛能够被快速地被ADO转化。在RNA或者DNA 脚手架的作用下,脂肪烃的产量确实分别得以提高1.8和8.8倍,但是总产量依然不尽如人意(只有27或者44mg/L)。
大肠杆菌具有内源的脂肪醛还原酶(aldehyde reductases,AHRs),可以将脂肪醛转化成脂肪醇分子。AHR也同样的以脂肪醛作为底物,因此看似好像与脂肪烃合成是竞争关系。但是,AHR的存在也同样可以促进脂肪醛转化的代谢流,并且能够降低脂肪醛的毒性。
发明内容
有鉴于此,本发明提供一种基因元件、表达载体及其应用。通过动态平衡关键中间代谢物脂肪醛的积累和转化来提高大肠杆菌脂肪烃合成的能力。为了达到这个目的,本专利首先通过协同表达脂肪醇合成过程,通过竞争路径帮助ADO平衡脂肪醛的合成与转化;随后将与脂肪醛动态平衡相关的脂肪酸合成、脂质降解和电子传递三个模块与脂肪烃合成模块相适配,最终实现工程大肠杆菌高产脂肪烃的人工合成。
为了实现上述发明目的,本发明提供以下技术方案:
本发明提供了aar基因和ado基因共表达在提高脂肪烃产量中的应用。
在本发明的一些具体实施方案中,aar基因和ado基因的表达强度为(0.60-1.0):(0.20-0.4)。
在本发明的一些具体实施方案中,还包括adhP基因与所述aar基因和ado基因共表达。
在本发明的一些具体实施方案中,aar基因、ado基因和adhP基因的表达强度为(0.60-1.0):(0.20-0.4):(0.10-0.60)。
在本发明的一些具体实施方案中,在AAR、ADO和AHR三者得到最好适配的三个菌中分别含有YX33(复制子:p15A;过表达的基因:PT7:aar-ado,PT7:aar-ado;RBS#:BBa_B0030;抗性:氯霉素),YX10(复制子:pBR322;过表达的基因:PT7:aar-ado;RBS#:BBa_B0030;抗性:氨苄)和YX45(复制子:pBR322;过表达的基因:PT7:ado-aar,PT7:aar;RBS#:Wild type;抗性:氨苄)三个质粒,在试管中发酵的脂肪烃总产量分别为:42.9mg/L、58.8mg/L和55.2mg/L。
在本发明的一些具体实施方案中,还包括过表达fabD基因、fab基因G或fabI基因中的一个或多个。
在本发明的一些具体实施方案中,不包括fabH基因、fabA基因、fabZ基因、fabB基因或accBCDA基因中的一个或多个基因的过表达。
在本发明的一些具体实施方案中,将脂肪酸合成路径中的每一个基因分别过表达,并与YX33,YX45或者YX10一起共转化到大肠杆菌E.coli BL21(DE3)菌株中,在试管中的发酵结果如图3A所示。无论是哪种启动子用于表达FAB中的基因,也无论是和哪个产烃质粒(YX33,YX45或者YX10)进行适配,总的来说,过表达fabD、fabG和fabI基因有助于提高烃的产出,而过表达fabH、fabA、fabZ、fabB和内源的accBCDA基因则会损害烃类物质的积累。当YX10与fabI基因一起过表达时,脂肪烃的产量可以达到81.8mg/L。
在本发明的另一些具体实施方案中,PT7和PTrc启动子分别用于表达FAB模块中的基因,并与YX33进行适配,其中PT7启动子的表达强度是PTrc的5倍。从图3B的发酵结果可以看出:1)当IPTG的诱导强度相同(1mM)时,在PT7启动子(高表达强度)表达下的FAB模块与YX33进行适配后,无论过表达FAB模块中的哪个基因,都严重损害了脂肪烃的产出。2)当采用相同启动子(PTrc)表达FAB模块中的基因时,分别采用了1mM,0.1mM和0.01mM三种浓度的IPTG进行诱导,发酵结果表明,当IPTG浓度过高(1mM)或者过低(0.01mM)时,无论过表达FAB模块中的哪个基因,脂肪烃的产出都比仅含有YX33质粒的对照菌株更低。只有当IPTG诱导浓度适中时,fabD、fabG和fabI基因的过表达才能够促进脂肪烃的产出。需要注意的是,低诱导浓度下(0.01mM),烃产出主要是因为YX33中AAR和ADO基因表达量不足所引起的。因此,在FAB模块与脂肪烃合成的适配中,FAB模块的表达强度不适宜过高。
在本发明的一些具体实施方案中,其还包括IPTG的诱导,所述IPTG的浓度为0.1mM。
在本发明的一些具体实施方案中,还包括aasS基因或者aasS-lipA基因共表达。
在本发明的一些具体实施方案中,当aasS或者aasS-lipA与YX45共表达的时候,脂肪烃的产量与只含有YX45质粒的菌株相比,分别被提高了30.2% 和21.1%。当aasS或者aasS-lipA与YX10共表达的时候,脂肪烃的产量与只含有YX10质粒的菌株相比,分别被提高了32.7%和32.1%。这些结果表明,表达解脂酶(Lipolytic enzymes,LipA)和脂酰-ACP合成酶(acyl-ACP synthetase,AasS),可以通过将代谢流从磷脂合成转向脂酰ACP前体供应,从而增加脂肪烃的产出。当YX10与aasS基因一起过表达时,脂肪烃的产量可以达到86.6mg/L。
在本发明的一些具体实施方案中,还包括来源于蓝细菌中的Se.Fd/FNR的过表达。
在本发明的一些具体实施方案中,不包括大肠杆菌内源的Ec.Fd/FNR的过表达。
在本发明的一些具体实施方案中,本发明通过过表达大肠杆菌内源(Ec.Fd/FNR)或者蓝细菌S.elongatus PCC7942中的(Se.Fd/FNR)铁氧还蛋白还原系统,与产烃过程进行适配。将还原力模块中各质粒分别与产烃质粒YX45和YX10一起转入E.coli BL21(DE3)中,在30℃诱导发酵40h,对发酵产物进行GC-MS检测,结果如图5所示。试管发酵结果表明,来源于蓝细菌中的还原系统(Se.Fd/FNR)与YX45或者YX10分别过表达后,脂肪烃的产出被提高到93.9或者101.7mg/L。而大肠杆菌内源的还原系统(Ec.Fd/FNR)则对烃类合成没有促进作用。当YX10与Se.Fd/Fnr基因一起过表达时,脂肪烃的产量可以达到101.7mg/L。
本发明还提供了所述应用获得的基因元件或表达载体。
在本发明的一些具体实施方案中,所述表达载体为大肠杆菌,其发酵的方式为分批补料发酵;所述分批补料发酵为在LB固体平板上30℃过夜培养,挑重组子单菌落接种于2mL LB培养基中30℃培养至OD为2.5~4,并按照1:100的比例转接于20mL M9培养基中,30℃培养至OD为2.5~4,并按照1:100的比例再次转接于800mL M9培养基中;当OD涨至2.5~4的时候,将培养液离心浓缩至50mL,并接种于发酵罐培养基中分批补料发酵;当OD升至15的时候,用10μM的IPTG进行诱导。
在本发明的一些具体实施方案中,所述发酵罐培养基包括6g/L NH4Cl,8.5g/L KH2PO4,1g/L MgSO4·7H2O,0.5g/L柠檬酸盐,0.07g/L CaCl2·2H2O, 15g/L甘油,5g/L酵母提取物,4mL的金属微量元素溶液以及4mL的10mg/L硫胺素(VB1)。金属微量元素溶液的浓度与实施例1中相同。
在本发明的一些具体实施方案中,所述分批补料发酵的补料溶液包括:2.47g/L MgSO4,500g/L甘油以及200g/L酵母提取物。
在本发明的一些具体实施方案中,含有YX10和Se.Fd/FNR质粒的大肠杆菌E.coli BL21(DE3)分批补料发酵结果如图6所示,诱导40.5h后,脂肪烃产量达到1.31g/L,OD值达到112.2。在发酵过程中,十五烷的比例在培养前34h从31.3%逐步提高到44.6%,并保持这个比例一直到发酵结束。而在培养34h后,也有少量的十五烯生成。
本发明还提供了所述的基因元件或表达载体在提高脂肪烃产量中的应用。
本专利提出一种提高脂肪烃微生物合成的新方法:通过动态平衡关键中间代谢物脂肪醛的积累和转化来提高大肠杆菌脂肪烃合成的能力。为了达到这个目的,本专利首先通过协同表达脂肪醇合成过程,通过竞争路径帮助ADO平衡脂肪醛的合成与转化;随后将与脂肪醛动态平衡相关的脂肪酸合成、脂质降解和电子传递三个模块与脂肪烃合成模块相适配,最终实现工程大肠杆菌高产脂肪烃的人工合成。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍。
图1示实施例1中降低竞争路径的脂肪醇合成对脂肪烃产出的负作用。(A)脂肪烃和脂肪醇微生物合成的代谢路径;(B)从大肠杆菌基因组中敲除主要负责十六醇合成的脂肪醛还原酶(AdhP),只能减少脂肪醇的产出,并不会促进脂肪烃的合成;(C)共同表达脂肪醇氧化酶(FAO)并不能通过将代谢流从脂肪醇引回脂肪醛的合成而增加脂肪烃的产出;
图2示实施例1中适当地保留竞争路径的脂肪醇合成对脂肪烃产出的正作用;(A)采用30个含有不同的aar和ado表达强度的质粒时,大肠杆菌E.coli BL21(DE3)的脂肪烃和脂肪醇的产出;其中(B)示(A)中结果的烃醇产出比;(C)不同工程菌株中脂肪醛的含量;(D)脂肪烃产出的三角图;颜 色代表脂肪烃的产量,每个基因诱导3h后的表达水平用实时定量PCR进行检测;*代表在这个适配过程中产量相对最高的菌株,并用于后续的优化;
图3示实施例2中内源脂肪酸合成与脂肪烃合成的适配结果;(A)脂肪酸合成过程中相关基因单独过表达,并分别与YX33、YX45和YX10进行适配;(B)脂肪酸合成过程中相关基因分别在不同的表达强度下与YX33进行适配;
图4示实施例3中内源脂质降解模块与脂肪烃合成的适配结果;通过过表达脂酰-ACP合成酶(acyl-ACP synthetase,AasS)和/或脂解酶(lipolytic enzymes,LipA)将碳代谢流从脂质往脂肪烃合成方向拉动;
图5示实施例4中还原力供给模块与脂肪烃合成的适配结果;通过过表达大肠杆菌内源(Ec.Fd/FNR)或者蓝细菌S.elongatus PCC7942中的(Se.Fd/FNR)铁氧还蛋白(ferredoxin,Fd)和铁氧还蛋白-NADP+还原酶(ferredoxin-NADP+reductase,FNR)来提高ADO的电子传递供给;*选作在实施例6中做进行分批补料发酵;
图6示实施例5中分批补料发酵的结果;图6A示实施例6中含YX10和Se.Fd/FNR的大肠杆菌E.coli BL21(DE3)分批补料发酵过程中生物量(OD600)、甘油消耗量、脂肪烃含量随时间变化曲线;图6B示实施例10中,分批补料发酵13小时、34小时、50.5小时后,发酵液中不同链长脂肪烃含量的比例。
具体实施方式
本发明公开了一种基因元件、表达载体及其应用,本领域技术人员可以借鉴本文内容,适当改进工艺参数实现。特别需要指出的是,所有类似的替换和改动对本领域技术人员来说是显而易见的,它们都被视为包括在本发明。本发明的方法及应用已经通过较佳实施例进行了描述,相关人员明显能在不脱离本发明内容、精神和范围内对本文所述的方法和应用进行改动或适当变更与组合,来实现和应用本发明技术。
本发明提供了aar基因和ado基因共表达在提高脂肪烃产量中的应用。在AAR、ADO和AHR三者得到最好适配的三个菌中分别含有YX33(复制子:p15A;过表达的基因:PT7:aar-ado,PT7:aar-ado;RBS#:BBa_B0030;抗性: 氯霉素),YX10(复制子:pBR322;过表达的基因:PT7:aar-ado;RBS#:BBa_B0030;抗性:氨苄)和YX45(复制子:pBR322;过表达的基因:PT7:ado-aar,PT7:aar;RBS#:Wild type;抗性:氨苄)三个质粒,在试管中发酵的脂肪烃总产量分别为:42.9mg/L、58.8mg/L和55.2mg/L。
本发明将脂肪酸合成路径中的每一个基因分别过表达,并与YX33,YX45或者YX10一起共转化到大肠杆菌E.coli BL21(DE3)菌株中,在试管中的发酵结果如图3A所示。无论是哪种启动子用于表达FAB中的基因,也无论是和哪个产烃质粒(YX33,YX45或者YX10)进行适配,总的来说,过表达fabD、fabG和fabI基因有助于提高烃的产出,而过表达fabH、fabA、fabZ、fabB和内源的accBCDA基因则会损害烃类物质的积累。当YX10与fabI基因一起过表达时,脂肪烃的产量可以达到81.8mg/L。
本发明将PT7和PTrc启动子分别用于表达FAB模块中的基因,并与YX33进行适配,其中PT7启动子的表达强度是PTrc的5倍。从图3B的发酵结果可以看出:1)当IPTG的诱导强度相同(1mM)时,在PT7启动子(高表达强度)表达下的FAB模块与YX33进行适配后,无论过表达FAB模块中的哪个基因,都严重损害了脂肪烃的产出。2)当采用相同启动子(PTrc)表达FAB模块中的基因时,分别采用了1mM,0.1mM和0.01mM三种浓度的IPTG进行诱导,发酵结果表明,当IPTG浓度过高(1mM)或者过低(0.01mM)时,无论过表达FAB模块中的哪个基因,脂肪烃的产出都比仅含有YX33质粒的对照菌株更低。只有当IPTG诱导浓度适中时,fabD、fabG和fabI基因的过表达才能够促进脂肪烃的产出。需要注意的是,低诱导浓度下(0.01mM),烃产出主要是因为YX33中AAR和ADO基因表达量不足所引起的。因此,在FAB模块与脂肪烃合成的适配中,FAB模块的表达强度不适宜过高。
当aasS或者aasS-lipA与YX45共表达的时候,脂肪烃的产量与只含有YX45质粒的菌株相比,分别被提高了30.2%和21.1%。当aasS或者aasS-lipA与YX10共表达的时候,脂肪烃的产量与只含有YX10质粒的菌株相比,分别被提高了32.7%和32.1%。这些结果表明,表达解脂酶(Lipolytic enzymes,LipA)和脂酰-ACP合成酶(acyl-ACP synthetase,AasS),可以通过将代谢流从磷脂合成转向脂酰ACP前体供应,从而增加脂肪烃的产出。当YX10与aasS基因一起过表达时,脂肪烃的产量可以达到86.6mg/L。
本发明通过过表达大肠杆菌内源(Ec.Fd/FNR)或者蓝细菌S.elongatus PCC7942中的(Se.Fd/FNR)铁氧还蛋白还原系统,与产烃过程进行适配。将还原力模块中各质粒分别与产烃质粒YX45和YX10一起转入E.coli BL21(DE3)中,在30℃诱导发酵40h,对发酵产物进行GC-MS检测,结果如图5所示。试管发酵结果表明,来源于蓝细菌中的还原系统(Se.Fd/FNR)与YX45或者YX10分别过表达后,脂肪烃的产出被提高到93.9或者101.7mg/L。而大肠杆菌内源的还原系统(Ec.Fd/FNR)则对烃类合成没有促进作用。当YX10与Se.Fd/Fnr基因一起过表达时,脂肪烃的产量可以达到101.7mg/L。
本发明通过含有YX10和Se.Fd/FNR质粒的大肠杆菌E.coli BL21(DE3)分批补料发酵结果如图6所示,诱导40.5h后,脂肪烃产量达到1.31g/L,OD值达到112.2。在发酵过程中,十五烷的比例在培养前34h从31.3%逐步提高到44.6%,并保持这个比例一直到发酵结束。而在培养34h后,也有少量的十五烯生成。
本发明提供的基因元件、表达载体及其应用中所用原料及试剂均可由市场购得。
下面的实施例是为了使本领域的技术人员能够更好地理解本发明,但并不对本发明作任何限制。
如在下列实施例中没有明确指出的操作,可以通过本领域技术人员常规的操作方法进行。
下面结合实施例,进一步阐述本发明:
实施例1:适当地利用竞争性AHR路径来提高脂肪烃产出。
实验材料:
1.基因
1)AAR:酰基-ACP还原酶,来源于聚球藻(Synechococcus elongatus PCC7942),蛋白序列为accession number:YP_400611,按照大肠杆菌进行密码子优化,优化之后的编码ADC7942的DNA分子具有aar所示的核苷酸序列,在GENEART合成基因。
2)ADO:醛基脱甲酰氧化酶,来源于聚球藻(Synechococcus elongatus PCC7942),蛋白序列为accession number:YP_400610,按照大 肠杆菌进行密码子优化,优化之后的编码ADO的DNA分子具有ado所示的核苷酸序列,在GENEART合成基因。
3)脂肪醇氧化酶(fatty alcohol oxidase,FAO),热带假丝酵母(Candida tropicalis,ATCC 20336)蛋白信息为:GenBank:AAS46878.1,Accession:AY538780.1,GI:44194456,按照大肠杆菌进行密码子优化,优化之后的编码FAO的DNA分子具有fao所示的核苷酸序列,在GENEART合成基因。
2.基因表达载体信息
1)质粒pBAD33购买自ATCC公司。
2)质粒pTrcHis2A购买自Invitrogen公司。
3)质粒pQE80L购买自Invitrogen公司。
4)质粒pACYCDuet-1购买自Novagen公司。
5)质粒pET21d购买自Novagen公司。
6)质粒pETDuet-1购买自Novagen公司。
3.基因操作所需试剂
1)TransStart FastPfu DNA聚合酶、TransFast Taq DNA聚合酶和dNTP(2.5mM each)购自北京全士金生物技术(TransGen Biotech)有限公司;
2)限制性内切酶和T4DNA连接酶购自Fermentas(Thermo Fisher,USA);
3)普通质粒小提试剂盒、琼脂糖凝胶DNA回收试剂盒、普通DNA产物纯化试剂盒购自天根生化科技(北京)有限公司。
4.内标和有机溶剂
十七烷和(128503-25G,99%)、十五醇(412228-5G,99%)和乙酸乙酯(产品编号3485,色谱溶剂99.7%)购自SIGMA-ALDICH公司(St.Louis,MO,USA);内标均用色谱纯无水乙醇(天津市天昊化工有限公司)溶解配制。
5.培养基配置所需试剂
1)异丙基硫代-β-D-半乳糖苷(IPTG)(纯度99%)购自天根生化科技(北京)有限公司;
2)酵母提取物购自OXOID(Thermo Fisher,USA);
3)琼脂粉、阿拉伯糖(分析纯)、Triton X100、和生物素购自北京鼎国昌盛生物技术有限责任公司;
4)LB和M9培养基中全部无机盐购自天津市北方天医化学试剂厂;
5)硫胺素(VB1)购自天津博美科生物技术有限公司;
6)甘油购自博欧特(天津)化工贸易有限公司
6.LB和M9培养基
LB液体培养基蛋白胨10g/L,酵母提取物5g/L,NaCl 10g/L,琼脂粉20g/L。M9矿物质培养基成分如下所示:12.8g/L的Na2HPO4·7H2O、3g/L KH2PO4、0.5g/L NaCl、2g/L NH4Cl、0.25g/L MgSO4·7H2O、14.7mg/L CaCl2、10mg/L硫胺素(VB1)、2g/L酵母提取物、30g/L甘油、0.1%(v/v)Triton-X100,以及1mL的如下浓度的微量元素:27g/L FeCl3·6H2O、2g/L ZnCl2·4H2O、2mg/L CaCl2、1.9mg/L CuSO4·5H2O、0.5mg/L H3BO3、2mg/L Na2MoO4·2H2O、100mL/L浓盐酸。pH值用Tris调节为7.25左右。
根据需要,添加的抗生素浓度为:氯霉素34μg/mL、氨苄霉素100μg/mL、卡纳霉素50μg/mL.
实验方法:
1.基因敲除过程
1)用于同源重组的引物序列如表1所示。采用对应上下引物将氯霉素从pKD3质粒中扩增下来,产物经琼脂糖凝胶电泳鉴定,并用琼脂糖凝胶DNA回收试剂盒回收PCR切胶产物;
2)将pKD46质粒转入大肠杆菌BL21(DE3)中,LB液体培养基中30℃过夜培养,活化后的菌体按照1:100的比例接种到含有10mmol/L L-阿拉伯糖的液体培养基中30℃培养,当菌体OD600长到0.5-0.6时冰上预冷10min,4℃,4000rpm离心5min(下同),随后用冰冷的10%甘油离心洗涤三次,浓缩100倍制成的电转感受态细胞,每管100μL,存放于-80℃冰箱待用;
3)采用电转化仪Electroporator 2170(Eppendorf,Germany)(0.1-cm chambers)将步骤1)中的PCR胶收产物(10-100ng)加入到感受态BL-46细胞中,1800V电击5-6ms后加入1mL无抗LB培养基于37℃,150rpm复苏3-4h,随后一半的细胞涂布在含有25μg/mL氯霉素的LB平板培养基中,剩余细胞室温放置过夜,如果24h后氯霉素平板中依然没有菌株生长,则将这些剩余细胞重新涂板。
4)对步骤3)中的平板挑单菌落,菌落PCR验证氯霉素是否将基因组中的对应基因替换下来。
5)将步骤4)中验证正确的转化子接到2mL氯霉素LB培养基中,43℃培养12h,将pKD46质粒缺失。划线后挑同一单菌落同时涂在氨苄和氯霉素的平板中,30℃培养24h,若同一单菌落在氯霉素平板中生长,在氨苄平板中不生长,则说明pKD46缺失完全。
6)将缺失过pKD46的转化子接菌制作电转感受态细胞,转入pCP20质粒,1mL无抗培养基30度复苏3-4小时后,吸出100μl接到2ml氨苄和氯霉素双抗培养基中30℃过夜培养,随后按照1:200的比例转接至无抗培养基中,43℃培养至稳定器后在无抗LB平板中划线,取单菌落在氨苄和氯霉素平板中分别划线验证染色体中氯霉素的弹出和pCP20质粒的丢失。并进行PCR和测序验证。
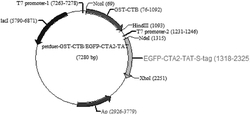
2.构建含有不同AAR,ADO和AdhP表达强度的质粒。每个质粒所包含的基因,核糖体结合位点(ribosome bind site,RBS),启动子,复制子,抗生素信息如表2所示。
3.将表2中的各质粒热激转化进入E.coli BL21(DE3)菌株中,并在LB固体平板上进行筛选。细胞均在30℃的培养箱中进行培养,固体及液体培养基中各抗生素的含量为氯霉素34μg/mL。
4.将转化了各质粒的E.coli BL21(DE3)菌株进行发酵。挑重组子单菌落接种于LB培养基中30℃过夜培养,并按照1:100的比例接种于5mL M9培养基中,在30℃,220rpm的摇床中进行进行发酵。当生物量长到OD600=1.0-1.2之间时,加入1mM的IPTG,诱导表达40h之后,提取产物进行检测。
5.产物的提取,具体做法如下:
1)取诱导后30℃发酵40h的培养基0.5mL,加入25mg/L的十八烷和 25mg/L十五醇作为内标;
2)加入0.5mL的乙酸乙酯,漩涡震荡5min,15000rpm离心2min;
3)吸取上层有机相,用0.22μm尼龙膜过滤。进样前将样品保存在-80℃冰箱中。
6.产物的检测,本实验中涉及的气质联用系统为Waters GCT Premier MICROMASS系统,其中包括:
1)Agilent 7683自动进样器
2)Agilent 6890气相色谱(GC,Agilent Technologies,USA)
3)飞行时间质谱仪(TOF-MS,Waters Corp.,USA)
4)J&W DB-5毛细管石英柱(30m length,I.D.0.25mm,Film 0.25μm,Agilent Technologies,USA)
GC条件如下:采用DB-5气相色谱柱,进样量为1μL,采用柱后分流技术,分流比为2:1。进样口温度为280℃,GC interface温度为280℃。高纯氦为载气,91Kpa恒压。色谱分离的升温程序如下:初始温度70℃,保持1min,分别以15℃·min-1、8℃·min-1、20℃·min-1的速度升到140℃、220℃、280℃,并在140℃和280℃分别保持1min和3min。
质谱条件如下:质谱电离方式为正离子模式的电轰击电离(EI+),其电离电压为70eV,源温保持在250℃。质谱的扫描范围为50-800m/z,扫描速度为2scan·s-1。
7.产物的定性和定量分析采用Masslynx软件(Version 4.1,Waters Corp.,USA)对GC-TOF/MS数据进行定性定量分析。采用NIST数据库(National Institute of Standard and Technology library,NIST,2005,Gaithersburg,MD)识别色谱峰,并用QuanLynx软件对各代谢物峰面积进行自动积分。通过各种物质的总离子流图的峰面积与同一张谱图上内标的峰面积的比值以获得产物的相对浓度值。
表1 实施例1中敲除所需引物
*下划虚线表示敲除所需的50-nt同源臂。起始和终止密码子用蓝色高亮标示。
表2 实施例1中的质粒信息
#核糖体结合位点(ribosome bind site,RBS)的序列和来源请见表2。
*(GCG)aar的RBS和基因起始序列为AGGAGGTACTAGATGGCGTTTGGC。GCG密码子被加到原始的第二个密码子TTT之前。(ala)ado的RBS和基因起始序列为AGGAGGTACTAGATGGCGCAGCTG.第二个密码子TTT被替换为GCG密码子。(下划线处为RBS序列,起始和终止密码子标注为蓝色)。
表3 RBS信息
下划线处为RBS序列,起始和终止密码子标注为蓝色
实验结果:
以脂酰-酰基运载蛋白(acyl carrier protein,ACP)为前体物的脂肪烃人工合成路径如图1A所示,其中脂酰ACP(Cn)被酰基-ACP还原酶(fatty acyl-ACP reductase,AAR)转化为脂肪醛(Cn),脂肪醛在醛基脱甲酰氧化酶(aldehyde deformylating oxygenase,ADO)的作用下氧化脱去羰基形成建一个碳的烷烃或者烯烃分子(Cn-1)。AAR和ADO两种酶都来源于蓝细菌(S.elongatus PCC7942),在大肠杆菌BL21(DE3)菌株内异源表达,因此作为外源脂肪烃合成模块。在这个人工合成体系中,脂肪烃的产物为:十五烷(pentadecane)和8-十七烯(8-heptadecene),分别由链长为C16和C18的脂酰-ACP经还原和脱甲酰氧化获得。
另一方面,脂肪醛还原酶(aldehyde reductase,AHR)也以中间代谢产物脂肪醛为前体物,将醛基还原成羟基,形成脂肪醇。大肠杆菌内源有数十种AHR,即使在没有人工构建脂肪醇合成路径的情况下,也一样能检测到脂肪醇的产出,因此脂肪醇合成是内源模块。在这数十种AHR中,AdhP蛋白主要 转化链长为C16的脂肪醛。十六醇也刚好是该人工合成体系中唯一检测到的脂肪醇产物,因此AdhP视为本研究中主要负责的AHR。
外源脂肪烃合成与内源脂肪醇合成模块同时使用AAR还原的脂肪醛作为前体物,因此理论上应该是互为竞争关系。但通过敲除基因组中adhP基因发现,虽然脂肪醇的合成确实能够减少70%,但是脂肪烃的合成也随之减少了30%(图1B),同时脂肪醛的积累从0提高到2.7-3.4mg/L。另外一方面,本研究表达了一种来源于在热带假丝酵母(Candida tropicalis,ATCC 20336)中的脂肪醇氧化酶(fatty alcohol oxidase,FAO),以期使碳代谢流从脂肪醇引流回中间代谢物脂肪醛。但是发酵结果表明,过表达了FAO之后,虽然脂肪醇产出确实得到阻遏,但是脂肪烃的产量也减少了一半,类似的,脂肪醛的积累也从1.0mg/L提高到了5.4mg/L。所有削弱脂肪醇合成的手段都会同时减少脂肪烃的产出。此外,烃的产量与脂肪醛中间代谢物的积累呈负相关。因此本专利认为,脂肪醛是AAR-ADO合成路径中的关键中间代谢物,内源脂肪醇合成途径可以通过提供中间代谢物的动态平衡,为原本为竞争关系的外源脂肪烃合成产生协同促进作用。
为了证实这个假设,也为了更好的适配两个内外源合成模块,本研究构建aar和ado表达强度各不相同的30个表达载体,其在大肠杆菌中发酵结果和脂肪烃/醇的比值分别如图2A和2B所示。将产出以及比值结果分为四个组,可以看到,脂肪烃产出最高的一组并不是分布在烃醇比值最高的位置。而是落在脂肪醇产出相对较高的III组。并且在第III组中,脂肪醛的积累也相对较少。在烃产出最高的YX10和YX45菌株中,脂肪醛的积累几乎为零,表明在高产烃的菌株中脂肪醛达到一个供给和消耗的动态平衡。,因此证实了上述假设。
此外,本研究还利用实时定量PCR的手段检测了aar,ado和adhP三个基因的转录水平,结果如图2所示。在这个重心坐标(barycentric coordinate)中,每个角代表标注在这个角处的基因的纯表达。而三角图(ternary plot)中每个点的位置,是由这个菌株中三个基因表达比例关系决定的,其颜色是由诱导发酵40h后的脂肪烃产量决定的。从这个三角图我们可以看出,脂肪烃的产量不仅是由aar和ado的表达量决定的,也同时由adhP的表达量决定。低烃产出(0-25mg/L)的菌株集中在重心坐标特别靠近或者特别远离adhP 的地方,也就是adhP表达量特别大或者特别小。而高脂肪烃产出的菌株大多集中在三角图中间靠下的位置(III),在这个区域中,aar:ado:adhP的表达强度比例大体为0.60-1.0:0.20-0.4:0.10-0.60(YX10附近)。这个转录结果证实了本研究的假设,即高产脂肪烃需要AHR的适量表达。
图2原始数据如表4所示:
表4
在AAR、ADO和AHR三者得到最好适配的三个菌中分别含有YX33,YX10和YX45三个质粒,在试管中发酵的脂肪烃总产量分别为:42.9mg/L、58.8mg/L和55.2mg/L。
实施例2:脂肪酸合成的适配
实验材料:
菌株信息、试剂和培养基等与实施例1中相同
实验方法:
1.YX33、YX10和YX45的构建方法和质粒信息与实施例1中相同。
2.过表达脂肪酸合成路径中各基因的质粒信息见表5。
3.将YX33、YX10和YX45分别与表5中的质粒热激转化进入E.coli BL21(DE3)菌中,并在LB固体平板上进行筛选。细胞均在30℃的培养箱中进行培养。当YX33、YX10和YX45与Trc-Fab系列基因共同转化进入E.coli BL21(DE3)菌株发酵时,IPTG浓度分成1mM,0.1mM和0.01mM三种浓度进行诱导。
4.将转化了各质粒的菌株进行发酵,,方法过程同实施例1。
5.产物的提取,方法过程同实施例1。
6.产物的检测,方法过程同实施例1。
表5 实施例3中的质粒信息
实验结果:
脂肪烃的合成以脂肪酸碳链延长为基础,因此与内源脂肪酸合成(fatty acid biosynthesis,FAB)模块息息相关。因此本专利针对上游脂肪酸合成模块与脂肪烃合成模块进行了适配性。我们将脂肪酸合成路径中的每一个基因分别过表达,并与YX33,YX45或者YX10一起共转化到大肠杆菌E.coli BL21(DE3)菌株中,在试管中的发酵结果如图3A所示。无论是哪种启动子用于表达FAB中的基因,也无论是和哪个产烃质粒(YX33,YX45或者YX10)进行适配,总的来说,过表达fabD、fabG和fabI基因有助于提高烃的产出,而过表达fabH、fabA、fabZ、fabB和内源的accBCDA基因则会损害烃类物质的积累。当YX10与fabI基因一起过表达时,脂肪烃的产量可以达到81.8mg/L,是本实施例中的最高产量。
另外,PT7和PTrc启动子分别用于表达FAB模块中的基因,并与YX33进行适配,其中PT7启动子的表达强度是PTrc的5倍。从图3B的发酵结果可以看出:1)当IPTG的诱导强度相同(1mM)时,在PT7启动子(高表达强度)表达下的FAB模块与YX33进行适配后,无论过表达FAB模块中的哪个基因,都严重损害了脂肪烃的产出。2)当采用相同启动子(PTrc)表达FAB模块中的基因时,分别采用了1mM,0.1mM和0.01mM三种浓度的IPTG进行诱导,发酵结果表明,当IPTG浓度过高(1mM)或者过低(0.01mM)时,无论过表达FAB模块中的哪个基因,脂肪烃的产出都比仅含有YX33质粒的对照菌株更低。只有当IPTG诱导浓度适中时,fabD、fabG和fabI基因的过表达才能够促进脂肪烃的产出。需要注意的是,低诱导浓度下(0.01mM),烃产出主要是因为YX33中AAR和ADO基因表达量不足所引起的。因此,在FAB模块与脂肪烃合成的适配中,FAB模块的表达强度不适宜过高,比如使用PTrc强度以下的启动子进行表达,IPTG诱导浓度低于等于0.1mM。
图3的原始数据见表6。
表6
实施例3:脂质降解的适配
实验材料:
菌株信息、试剂和培养基等与实施例1中相同。
实验方法:
1.YX33、YX10和YX45的构建方法和质粒信息与实施例1中相同。
2.脂质降解系列质粒信息见表7。
3.将YX33、YX10和YX45分别与表7中的质粒热激转化进入E.coli BL21(DE3)菌中,并在LB固体平板上进行筛选。细胞均在30℃的培养箱中进行培养。当YX33、YX10和YX45与Trc-Fab系列基因共同转化进入E.coli BL21(DE3)菌株发酵时,IPTG浓度分成1mM,0.1mM和0.01mM三种浓度进行诱导。
4.将转化了各质粒的菌株进行发酵,,方法过程同实施例1。
5.产物的提取,方法过程同实施例1。
6.产物的检测,方法过程同实施例1。
表7 实施例4中的质粒信息
实验结果:
另外一方面,脂酰-ACP以及自由脂肪酸还可以参与细胞膜的磷脂合成,因此脂肪烃合成也与脂质降解模块密切关联。解脂酶(Lipolytic enzymes,LipA)能够将膜上的磷脂水解成自由脂肪酸,再经脂酰-ACP合成酶(acyl-ACP synthetase,AasS)的催化,被水解的自由脂肪酸又可以被活化连接到与酰基运载蛋白(ACP)上,增加脂酰ACP前体的供给。本专利通过过表达集胞藻Synechocystis sp.PCC 6803中的lipA基因和哈氏弧菌Vibrio harveyi B392中的aasS基因,将磷脂降解与脂肪烃合成进行适配。这些基因都进行了大肠杆菌密码子优化。将各质粒转入E.coli BL21(DE3)中,在30℃诱导发酵40h,对发酵产物进行GC-MS检测,结果如图4所示。当aasS或者aasS-lipA与YX45共表达的时候,脂肪烃的产量与只含有YX45质粒的菌株相比,分别被提高了30.2%和21.1%。当aasS或者aasS-lipA与YX10共表达的时候,脂肪烃的产量与只含有YX10质粒的菌株相比,分别被提高了32.7%和32.1%。这些结 果表明,表达解脂酶(Lipolytic enzymes,LipA)和脂酰-ACP合成酶(acyl-ACP synthetase,AasS),可以通过将代谢流从磷脂合成转向脂酰ACP前体供应,从而增加脂肪烃的产出。当YX10与aasS基因一起过表达时,脂肪烃的产量可以达到86.6mg/L,是本实施例中的最高产量。
图4的原始数据如表8所示。
表8
实施例4:还原力的适配
实验材料:
菌株信息、基因信息、试剂和培养基等与实施例1中相同
实验方法:
1.YX33、YX10和YX45的构建方法和质粒信息与实施例1中相同。
2.还原力供给系列质粒信息见表9。
3.将YX33、YX10和YX45分别与表9中的质粒热激转化进入E.coli BL21(DE3)菌中,并在LB固体平板上进行筛选。细胞均在30℃的培养箱中进行培养。当YX33、YX10和YX45与Trc-Fab系列基因共同转化进入E.coli BL21(DE3)菌株发酵时,IPTG浓度分成1mM,0.1mM和0.01mM三种浓度进行诱导。
4.将转化了各质粒的菌株进行发酵,,方法过程同实施例1。
5.产物的提取,方法过程同实施例1。
6.产物的检测,方法过程同实施例1。
表9 实施例5中的质粒信息
实验结果:
ADO发挥活性需要辅助还原系统为其提供电子供给。铁氧还蛋白(ferredoxin,Fd)和铁氧还蛋白-NADP+还原酶(ferredoxin-NADP+reductase,FNR)是生物还原系统中被应用的最为广泛的。本专利通过过表达大肠杆菌内源(Ec.Fd/FNR)或者蓝细菌S.elongatus PCC7942中的(Se.Fd/FNR)铁氧还蛋白还原系统,与产烃过程进行适配。将还原力模块中各质粒分别与产烃质粒YX45和YX10一起转入E.coli BL21(DE3)中,在30℃诱导发酵40h,对发酵产物进行GC-MS检测,结果如图5所示。试管发酵结果表明,来源于蓝细菌中的还原系统(Se.Fd/FNR)与YX45或者YX10分别过表达后,脂肪烃的产出被提高到93.9或者101.7mg/L。而大肠杆菌内源的还原系统(Ec.Fd/FNR)则对烃类合成没有促进作用。当YX10与Se.Fd/Fnr基因一起过表达时,脂肪烃的产量可以达到101.7mg/L,是本实施例中的最高产量。
图5原始数据如表10所示。
表10
实施例5:分批补料发酵
实验材料:
菌株信息、试剂和试管发酵培养基等与实施例1中相同。
发酵罐培养基:6g/L NH4Cl,8.5g/L KH2PO4,1g/L MgSO4·7H2O,0.5g/L柠檬酸盐,0.07g/L CaCl2·2H2O,15g/L甘油,5g/L酵母提取物,4mL的金属微量元素溶液以及4mL的10mg/L硫胺素(VB1)。金属微量元素溶液的浓度与实施例1中相同。
补料溶液中物质和浓度:2.47g/L MgSO4,500g/L甘油以及200g/L酵母提取物。
实验方法:
1.YX10的构建方法和质粒信息与实施例1中相同。
2.Se.Fd/FNR的构建方法和质粒信息与实施例4中相同。
3.将YX10和Se.Fd/FNR质粒热激转化进入E.coli BL21(DE3)菌株中,并在LB固体平板上30℃过夜培养,挑重组子单菌落接种于2mL LB培养基中30℃培养至OD为2.5-4,并按照1:100的比例转接于20mL M9培养基中,30℃培养至OD为2.5-4,并按照1:100的比例再次转接于800mL M9培养基中。当OD涨至2.5-4的时候,将培养液离心浓缩至50mL,并接种与2.5L的发酵罐中进行分批补料发酵。当OD升至15的时候,用10μM的IPTG进行诱导。每隔4h取样,每次取15mL用于细胞密度、甘油、乙酸、脂肪醇浓度的分析。固体及液体培养基中各抗生素的含量为氯霉素34μg/mL、氨苄100μg/mL。
4.细胞密度采用TU-1810紫外可见分光光度计(北京普析通用仪器有限公司),在波长600的条件下进行测量。
5.甘油浓度的测量:取1mL发酵液12,000rpm离心10min,取上清用0.22μm滤膜过滤,根据情况进行稀释,或者直接进样到HPLC中进行分离检测。HPLC为Waters e2695,检测器为2414RI示差检测器,色谱柱为Aminex HPX-87H column(BioRad,CA),柱温保持在65℃,流动相为5mM的稀硫酸水溶液,流速为0.6mL/min。
6.产物的提取,方法过程同实施例1。
7.产物的检测,方法过程同实施例1。
实验结果:
含有YX10和Se.Fd/FNR质粒的大肠杆菌E.coli BL21(DE3)分批补料发酵结果如图6所示,诱导40.5h后,脂肪烃产量达到1.31g/L,OD值达到112.2。在发酵过程中,十五烷的比例在培养前34h从31.3%逐步提高到44.6%,并保持这个比例一直到发酵结束。而在培养34h后,也有少量的十五烯生成。
图6的原始数据如表11所示。
表11
以上对本发明所提供的基因、其编码的蛋白质及用途、基因元件、利用竞争路径与模块化代谢流适配提高大肠杆菌脂肪烃微生物合成的方法进行了详细介绍。本文应用了具体个例对本发明的原理及实施方式进行了阐述,以上实施例的说明只是用于帮助理解本发明的方法及其核心思想。应当指出,对于本技术领域技术人员来说,在不脱离本发明原理的前提下,还可以对本发明进行若干改进和修饰,这些改进和修饰也落入本发明权利要求的保护范围内。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
基因元件、表达载体及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0